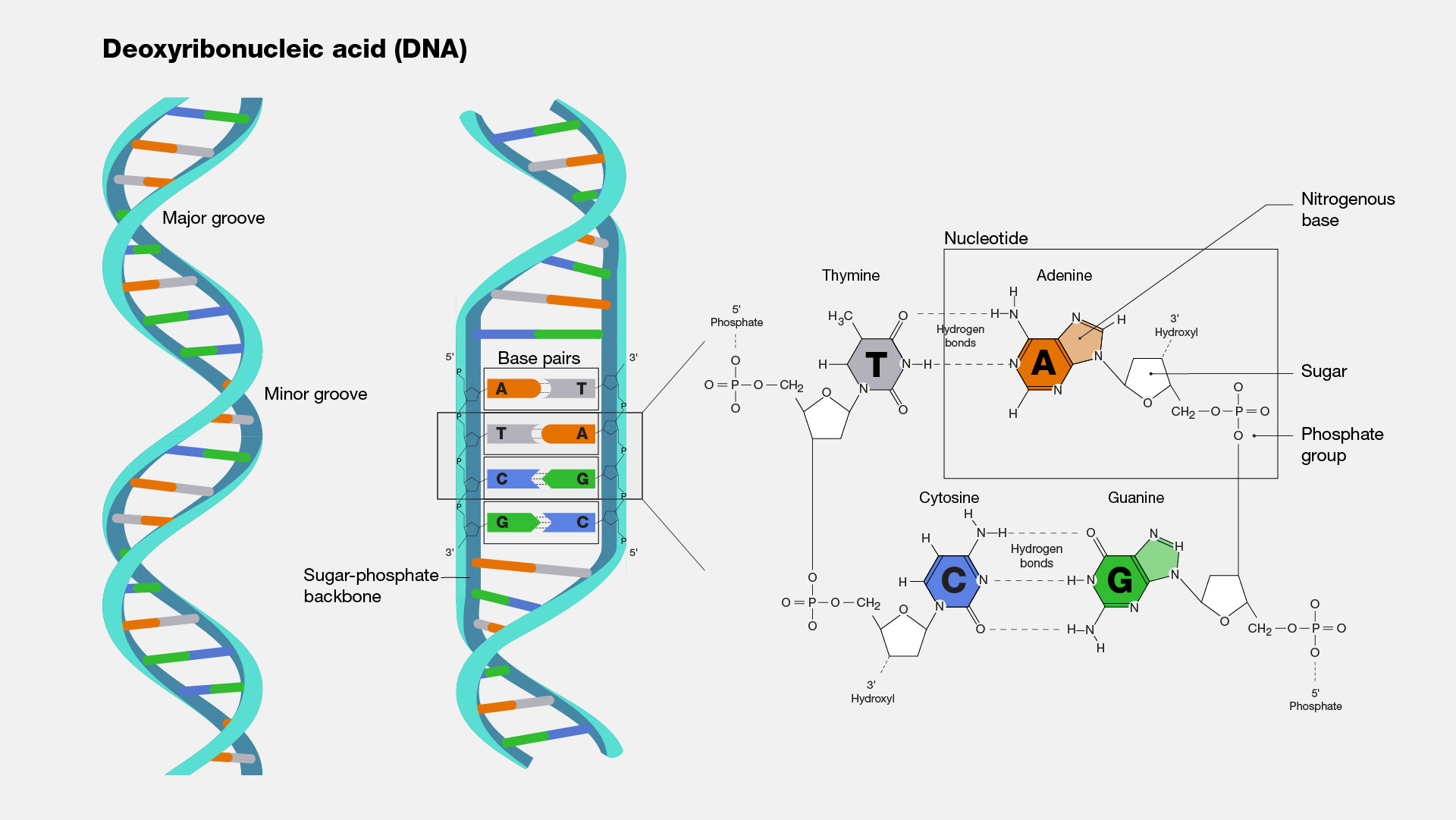
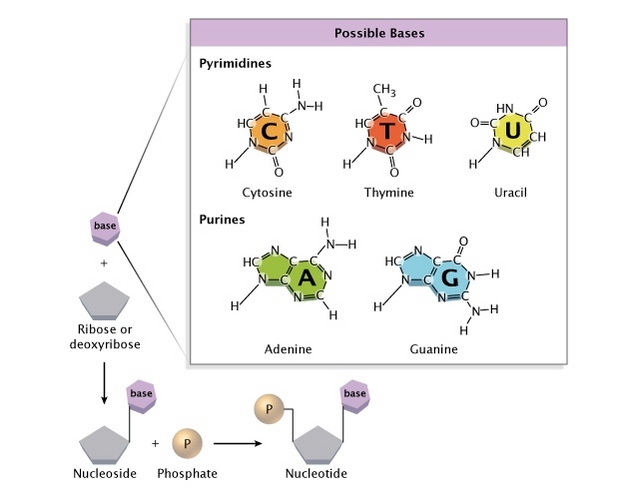
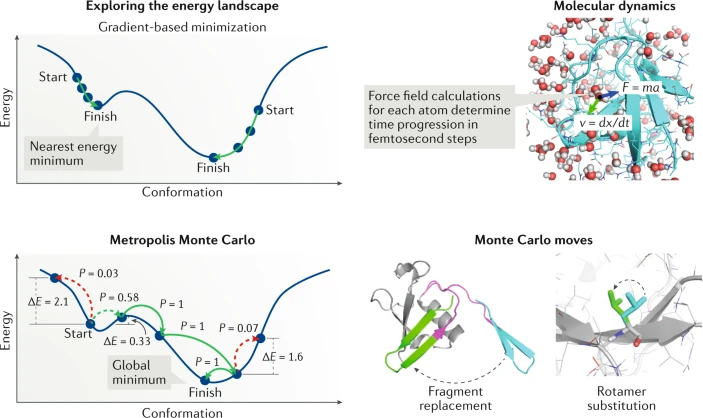
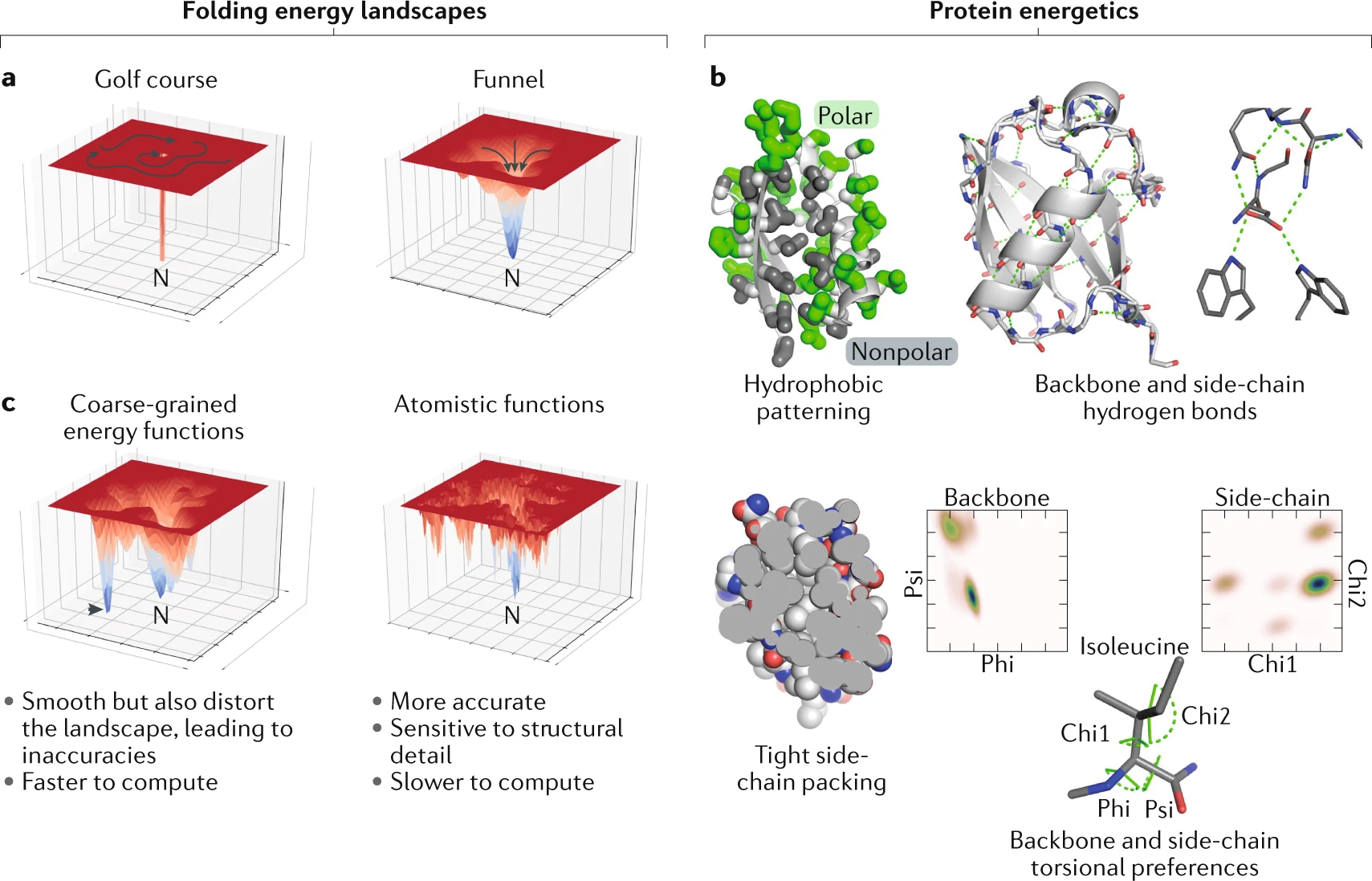
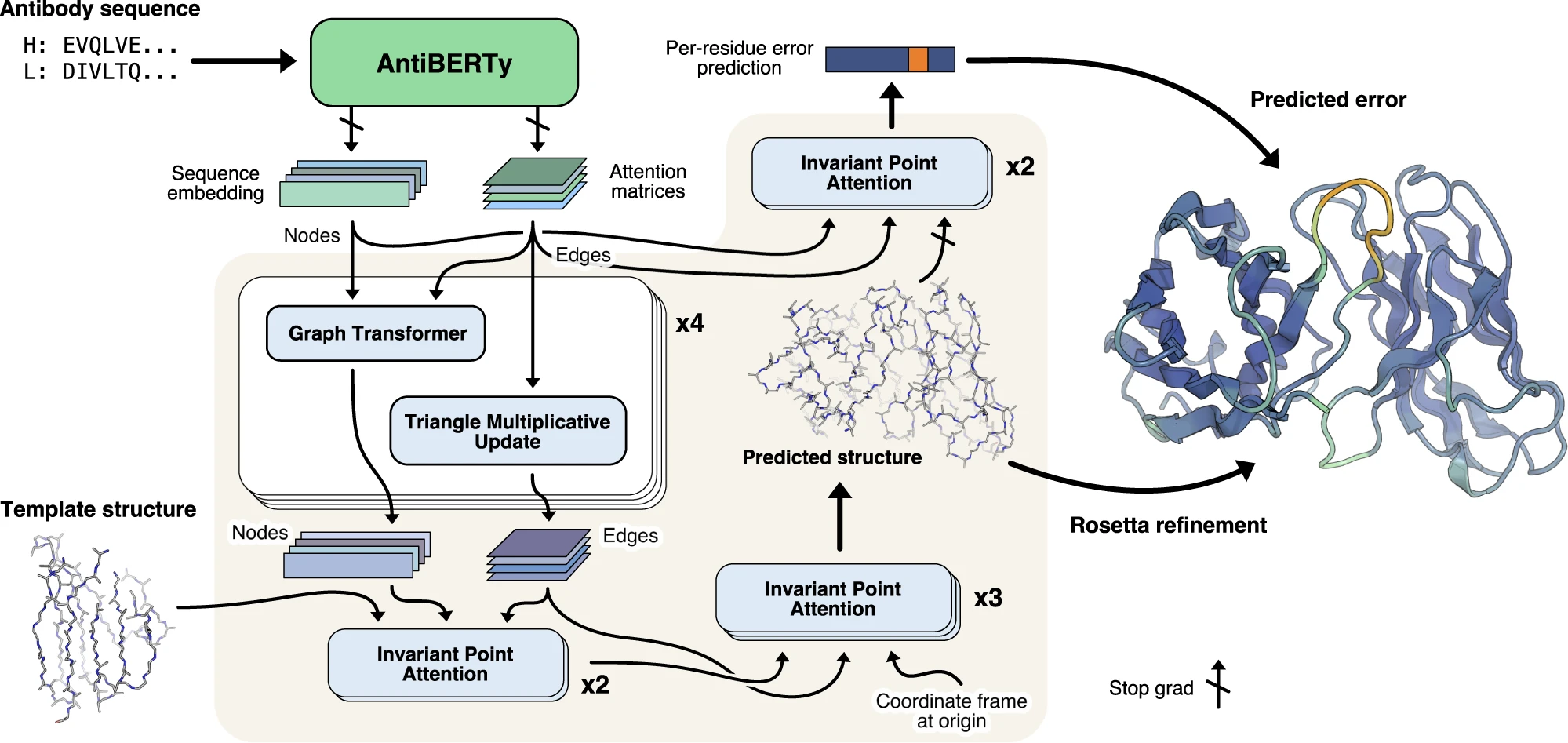
단백질은 하나 또는 그 이상의 폴리펩타이드가 꼬이거나 접혀서 생성된 독특한 3차원 구조(입체 구조)를 가진다. 단백질 구조는 보통 자발적으로 생성된다. 단백질이 세포에서 합성되면서 아미노산 서열에 따라 사슬의 부분 사이에 다양한 형태의 결합이 형성된다. 단백질의 독특한 형태는 다른 분자를 인식하고 결합할 수 있도록 한다. 구형 단백질은 대략 구 모양을 띠며 섬유성 단백질은 긴 섬유 모양이다.
1차 구조(primary structure)는 유전적으로 암호화된 단백질 내의 아미노산 서열이다.
2차 구조(secondary structure)는 한 펩타이드 결합의 산소와 다른 펩타이드 결합의 질소에 부착한, 부분적으로 양전하를 띠는, 수소 사이의 수소 결합으로 안정화되는, 폴리펩타이드 골격의 꼬임 또는 접힘 부위를 포함한다. α-나선(α-helix)은 매 4번째 아미노산 사이의 수소 결합으로 형성된 나선이다. β 병풍구조(β pleated sheet)는 서로 평행하게 놓인 폴리펩타이드 골격 부위를 따라 반복되는 수소 결합으로 연결되어 있다.
3차 구조(tertiary structure)는 아미노산의 다양한 곁사슬 (R기) 사이의 상호작용으로 인해 생긴 단백질의 3차원 구조이다. 아래와 같은 여러 화학적 상호작용이 안정되고 독특한 단백질 모양을 이루도록 한다: 물에 의한 반발력에 의해 분자의 중심에 모여 덩어리를 이룬 비극성 곁사슬 사이의 소수성 상호작용(hydrophobic interaction), 비극성 곁사슬을 따라 존재하는 반데르발스 상호작용, 극성 곁사슬 사이의 수소 결합 및 음전하를 띤 곁사슬과 양전하를 띤 곁사슬 사이의 이온 결합. 이황화 결합(disulfide bridge)이라 불리는 강한 공유 결합이 폴리펩타이드가 접히면서 가까워진 시스테인의 황화수소기 사이에서 일어나기도 한다.
4차 구조(quaternary structure)는 둘 이상의 폴리펩타이드로 이루어진 단백질에서 일어난다. 각 폴리펩타이드 소단위는 구조적으로 정확하게 배열을 이루면서 연결되어 기능성 단백질을 형성한다.
운동은 체력을 향상시키고 건강을 유지하는 데 있어 적극적인 역할을 하는 것으로 오랫동안 알려져 왔습니다. 규칙적인 적당한 강도의 운동은 인간 건강의 모든 측면을 개선하며 다양한 질병에 대한 예방 및 치료 전략으로 널리 받아들여지고 있습니다. 운동은 유기체, 조직, 세포 및 분자 수준에서 항상성을 유지하고 복원하여 결과적으로 다양한 병리학적 상태로부터 보호하는 긍정적인 생리학적 적응을 자극한다는 것이 잘 문서화되어 있습니다. 여기에서는 중간 강도 운동이 장벽의 완전성, 국소적 교란의 억제, 재활용 및 전환, 회로 통합, 리듬 진동, 항상성 탄력성, 호르몬 조절, 회복 및 재생을 포함하여 건강의 주요 특징에 어떻게 영향을 미치는지 주로 요약합니다. 또한, 우리는 운동에 대한 반응으로 유익한 적응을 담당하는 메커니즘에 대한 현재의 이해를 요약합니다. 이 리뷰는 중간 강도의 운동이 건강을 유지하고 다른 건강 중재에 적용할 수 있는 창을 여는 중요한 생물학적 메커니즘에 대한 포괄적인 요약을 제공하는 것을 목표로 했습니다. 우리는 이 분야에 대한 지속적인 조사를 통해 중간 강도 운동의 긍정적인 역할과 관련된 과정에 대한 이해를 더욱 높이고 삶의 질을 향상시키는 새로운 치료법을 찾는 데 더 가까워지기를 바랍니다.
소개
현대화로 인해 인구 수명이 연장되었지만, 비만, 고혈압, 제2형 당뇨병, 암 등 비전염성 질병도 지속적으로 증가했습니다. 비전염성 질병은 특정 국가에서 사망의 80% 이상을 초래하기 때문에 이제 전 세계적으로 “제1의 살인자”로 간주됩니다. 비전염성 질병의 유병률은 적어도 부분적으로는 신체 활동이나 운동이 부족하기 때문일 수 있습니다. 세계보건기구(WHO)의 데이터에 따르면, 2016년에 전 세계 성인의 4분의 1 이상이 신체적으로 활동적이지 않았습니다. 신체 활동 부족이라는 세계적인 유행병은 공중 보건의 우선순위가 되어야 합니다. 건강한 생활 방식은 전체 사망률의 위험을 현저히 낮추고 기대 수명을 연장시키는 것과 관련이 있다는 것은 잘 알려져 있습니다. 건강하고 적절한 식이 패턴과 함께 운동은 비만과 관련된 만성 대사 질환 및 염증성 질환의 위험을 줄이기 위한 유망한 전략을 나타냅니다. 일반적으로 신체활동이란 걷기, 육체 노동, 집안일 등 에너지가 있어야 하는 모든 움직임으로 정의됩니다. 이에 반해 운동은 체력 향상을 목표로 계획되고 구성된 신체 활동의 루틴을 의미합니다. 이 리뷰에서 우리는 최대 산소 섭취량 (VO2max )이 70% 미만인 정기적인 중간 강도의 운동을 지칭하기 위해 “규칙적인 운동”이라는 용어를 사용합니다.
어린이와 성인은 앉아 있는 시간을 제한해야 하며, 이는 모든 원인으로 인한 사망, 심혈관 질환, 암, 제2형 당뇨병 발병 등 건강에 좋지 않은 결과와 관련이 있습니다. 앉아서 생활하는 행동을 모든 강도(빛의 강도 포함)의 신체 활동으로 대체하는 것은 건강에 유익합니다. 상당한 건강상의 이점을 위해 세계보건기구(WHO)는 성인이 일주일 내내 최소 150~300분의 중강도 또는 75~150분의 고강도 유산소 신체 활동을 수행하거나 중강도와 고강도 유산소 신체 활동을 이에 상응하는 조합으로 수행해야 한다고 권장합니다. 성인은 일주일 내내 중강도 유산소 신체활동을 300분 이상으로 늘리거나, 격렬한 유산소 신체활동을 150분 이상 수행하거나, 중강도와 고강도 유산소 신체활동을 동등한 조합으로 수행하도록 권장됩니다. 성인은 또한 모든 주요 근육 그룹을 포함하는 근육 강화 활동을 일주일에 2일 이상 중간 강도 이상의 강도로 수행하는 것이 좋습니다. 유산소 활동 다음으로 65세 이상 성인은 기능적 균형과 근력 운동을 포함한 복합 신체 활동을 중강도 이상의 강도로 일주일에 3일 이상 하는 것이 좋습니다.
임신 전 30시간의 대사 등가 작업은 임신성 당뇨병의 상대적 위험을 12% 감소시키는 것과 관련이 있고, 임신 전과 임신 중 주당 7시간의 신체 활동은 30% 감소와 관련이 있다는 증거가 있습니다. 위험은 각각 37% 감소했습니다. 또한, 주당 500회 대사량에 해당하는 유산소 신체 활동(중등도 신체 활동 150분 또는 고강도 신체 활동 75분에 해당)은 전체 원인으로 인한 사망 위험을 14% 감소시키는 것으로 나타났습니다. 심혈관 질환 환자는 건강한 성인의 경우 위험이 7% 감소합니다. 그리고 5~17세 어린이와 청소년은 하루 평균 60분 동안 중등도에서 격렬한 강도의 신체 활동을 수행했는데, 이는 심폐 건강, 근력 건강, 뼈 건강 및 심대사 건강을 포함한 여러 가지 유익한 건강 결과와 관련이 있었습니다.
많은 연구에서 규칙적인 운동이 심혈관 질환의 위험을 줄이고 비만, 제 2형 당뇨병, 다발성 경화증, 뇌졸중, 노화 관련 근육 감소증 및 일부 유형의 암 등 다양한 기타 병리를 앓고 있는 환자의 건강 결과를 개선하는 데 중요한 역할을 한다는 개념을 뒷받침합니다. 역학 연구에 따르면 심혈관 질환 유무와 관계없이 신체 활동 수준이 높을수록 사망 위험이 낮아지지만, 사망률과 관련하여 신체 활동의 이점은 더 큰 것으로 나타납니다. 심혈관 질환이 있는 사람과 심혈관 질환이 없는 사람의 비교. 더욱이, 규칙적인 신체 활동이나 운동은 일반적으로 건강에 해로운 결과(예: 장애 수준, 사망률)의 위험 감소 및 더 나은 생존과 관련이 있습니다. 운동은 낮은 신체 활동과 관련된 만성 질환과 복합 질병을 부분적으로 역전시킬 수 있으며 삶의 질을 향상시키기 위한 예방적 접근 방식으로 사용될 수 있습니다. 또한, 더 많은 양과 더 높은 강도의 신체 활동뿐만 아니라 다양한 유형의 신체 활동(예: 유산소 운동, 근육 및 뼈 강화 활동)은 여러 가지 유익한 건강 결과(예: 심폐 건강, 근육 강화 활동)와 관련이 있습니다. 5~17세 어린이 및 청소년을 위한 피트니스, 뼈 건강 및 심장 대사 건강).
근력, 지구력, 균형, 유연성, 협응력과 같은 규칙적인 중간 강도 유형의 운동은 인간 건강의 모든 측면에 유익하며 심혈관 질환(예: 심근병증, 심장병, 심장 질환)을 비롯한 다양한 질병에 대한 치료 및 예방 전략으로 널리 받아들여지고 있습니다. (허혈/ 재관류 손상, 심부전), 대사질환 (예: 고지혈증, 대사증후군, 제2형 당뇨병), 신경계 질환 (예: 파킨슨 병, 다발성 경화증), 폐질환 등. 그러나 신체 운동이 최소 강도 수준을 약화시키는 경우 신체 항상성에 큰 변화가 예상되지 않습니다. 훈련을 받지 않은 개인의 경우 갑자기 격렬한 강도의 운동을 시작하면 심혈관 질환이 발생할 수 있으며 장기간 강렬한 운동을 하면 경색의 발생률이 높아질 수 있습니다. 따라서 운동의 강도와 방식은 건강에 긍정적인 영향을 미치는 데 매우 중요합니다.
운동의 생물학은 복잡하며 여러 기관 시스템의 적응 반응을 포함합니다. 움직임과 신체 활동은 에너지 수요를 충족시키기 위해 골격근을 전신 조절자로 바꾸는 진화적 생존 이점입니다. 운동은 심혈관, 호흡기, 근골격계 뿐만 아니라 면역계와 내분비계에도 영향을 미치는 역동적인 에너지 소모 활동입니다. 운동의 효능에 대한 광범위한 조사에도 불구하고 운동의 이점에 대한 기본 메커니즘은 아직 파악하기 어렵습니다. 운동에 대한 심리사회적, 생물학적 반응을 요약한 논문이 꽤 많지만, 대부분은 특정 질병이나 장기 시스템에 초점을 맞추고 있습니다. 우리가 아는 한, 유기체의 전반적인 “조직”과 기본 메커니즘의 관점에서 운동으로 인한 변화를 보고하는 연구는 거의 없습니다.
최근 논문에서 이 연구의 공동 저자 중 2명은 건강의 8가지 특징을 제안했습니다: 장벽의 완전성, 국소적 교란의 억제, 재활용 및 회전율, 회로 통합, 리듬 진동, 항상성 탄력성, 호르몬 조절 및 회복 그리고 재생. 더 많은 조사가 필요하지만, 운동이 이러한 특징의 대부분에 영향을 미칠 수 있다는 증거가 풍부 합니다( 그림 1 ). 이 리뷰는 운동이 건강에 유익한 효과를 가져오는 중요한 생물학적 변화에 대한 포괄적인 요약을 제공하는 것을 목표로 했습니다. 또한, 이 리뷰에서는 운동 중재가 질병, 특히 연령 관련 질병(알츠하이머병, 파킨슨병), 제2형 당뇨병, 심혈관 질환(심근병증, 심장 허혈/재관류 손상, 심부전)을 예방하는 데 효과적인 메커니즘을 요약합니다. 및 특정 암(유방암 및 대장암). 더 나은 독해를 위해 동물 연구의 운동 프로토콜을 자세히 설명하는 보충 표 1, 인간 연구의 운동 프로토콜을 자세히 설명하는 표 1, 본 리뷰에서 인용된 무작위 대조 시험을 기반으로 한 증거를 요약한 표 2를 포함했습니다.
구체적인 내용은 원문에서 확인하세요.
운동은 장벽의 무결성을 보호합니다
운동은 국소 항상성 유지에 도움이 됩니다
재활용 및 회전율에 있어서 운동의 이점
다양한 회로에 대한 운동의 효과
리듬 진동에 대한 운동의 효과
항상성 회복력에 있어서 운동의 이점
호르몬 조절에 있어 운동의 이점
운동이 회복과 재생에 미치는 영향
운동이 건강에 미치는 통합적 효과
결론 및 전망
운동의 유익한 효과는 대사 및 심폐 건강 관련 결과의 개선에 대해 잘 문서화되어 있습니다. 규칙적인 운동은 본질적으로 특정 동반 질환이 있는 환자에게 비약리학적 폴립 제로 간주됩니다. 치료 운동은 비만, 제2형 당뇨병, 심혈관 질환, 노화 관련 근육 위축 및 특정 암과 같은 질병을 예방, 관리 및 치료하기 위해 채택될 수 있습니다. 그러나 운동 프로토콜은 연구마다 다를 수 있으므로 이러한 발견을 해석할 때는 주의해야 합니다.
운동은 자유로운 생활을 하는 개인을 정확하게 평가하기 어려운 복잡하고 다차원적인 과정입니다. 운동을 정량화하는 데 적용되는 방법의 일관성 부족은 데이터 분석의 한계이며 실험 결과를 임상 실습으로 전환하는 데 방해가 될 수 있습니다. 따라서 운동과 관련된 향후 작업은 재현성이 뛰어나고 엄격하게 설계되고 잘 보고된 시험을 기반으로 수행되어야 합니다.
본 리뷰에서 우리는 생물학적 원인의 관점에서 운동의 건강상의 이점을 요약하려고 시도했습니다. 지면의 제약으로 인해 개별 주제별로 종합적인 리뷰를 제공하기는 어렵습니다. 그러나 지금까지 논의된 주제와 특징은 운동 관련 연구에 대한 기계적 관점을 제공할 수 있습니다. 운동의 생물학은 복잡하므로 각 주제와 특성에 대한 운동의 효과를 설명하기 위해 예를 나열했으며, 이는 운동으로 인한 적응을 포괄적으로 이해하는 데 도움이 될 수 있습니다.
현재의 Mendelian 무작위화 연구와 섹션 7.2 의 전통적인 관찰 연구 사이의 상충되는 결과와 관련하여 미래의 연구자들은 운동의 생물학적 경로를 밝히기 위해 더욱 기계적인 연구를 수행해야 합니다. 게다가 운동으로 자극된 기관간 의사소통에 대해서는 아직 연구가 덜 되어 있습니다. 운동과 관련된 기관간 의사소통과 그 기본 메커니즘에 대한 더 자세한 연구는 운동 반응 분자를 식별하는 데 도움이 될 수 있습니다. 규칙적인 운동은 의심할 여지 없이 전반적인 건강에 유익합니다. 그러나 운동의 다양한 건강상의 이점을 뒷받침하는 메커니즘은 복잡하며 세포 소기관(예: 미토콘드리아, 핵, ER)의 적응을 탐구하는 데 전념하는 노력은 운동의 긍정적인 효과를 이해하는 데 도움이 될 수 있습니다.
운동의 유익한 효과를 뒷받침하는 메커니즘에 대한 추가 조사는 운동 중재의 설계를 개선하고 치료 효과를 극대화하는 데 도움이 될 것입니다. 그러므로 건강 증진을 위한 운동 방법의 수정을 지향하는 잠재적인 후속 개입의 효과를 더 잘 이해하고 사람의 건강을 향상할 수 있는 특정 분자를 연구하기 위해서는 운동의 과학적 기초에 대한 사전 심층 지식이 필요합니다. 우리는 이 분야에 대한 지속적인 조사가 운동의 유익한 역할과 관련된 과정에 대한 이해를 높이고 삶의 질을 향상하기 위한 새로운 치료법의 식별을 촉진할 수 있기를 바랍니다.
암의 특징은 인간 종양의 다단계 발달 중에 획득된 6가지 생물학적 능력으로 구성됩니다. 특징은 종양성 질환의 복잡성을 합리화하기 위한 조직 원칙을 구성합니다. 여기에는 증식 신호 유지, 성장 억제 인자 회피, 세포 사멸 저항, 복제 불멸 활성화, 혈관 신생 유도, 침입 및 전이 활성화가 포함됩니다. 이러한 특징의 기본은 게놈의 획득을 촉진하는 유전적 다양성을 생성하는 게놈 불안정성과 여러 특징 기능을 촉진하는 염증입니다. 지난 10년간의 개념적 진보로 인해 이 목록에 잠재적인 일반성의 두 가지 새로운 특징, 즉 에너지 대사 재프로그래밍과 면역 파괴 회피가 추가되었습니다. 암세포 외에도 종양은 또 다른 차원의 복잡성을 나타냅니다. 여기에는 “종양 미세환경”을 생성하여 특징적인 특성을 획득하는 데 기여하는 표면적으로는 정상인 모집된 세포의 레퍼토리가 포함되어 있습니다. 이러한 개념의 광범위한 적용 가능성에 대한 인식은 인간 암을 치료하는 새로운 수단의 개발에 점점 더 영향을 미칠 것입니다.
본문
소개
우리는 암의 6가지 특징이 함께 종양성 질환의 놀라운 다양성을 이해하기 위한 논리적 틀을 제공하는 구성 원리를 구성한다고 제안했습니다. (하나한과 와인버그, 2000). 우리의 논의에는 정상 세포가 종양성 상태로 점진적으로 진화함에 따라 이러한 특징적인 능력을 연속적으로 획득하고 인간 종양 발병의 다단계 과정이 초기 암세포가 이러한 특성을 획득해야 한다는 필요성에 의해 합리화될 수 있다는 개념이 내포되어 있습니다. 종양을 유발하고 궁극적으로 악성이 될 수 있습니다.
우리는 종양이 증식하는 암세포의 섬 덩어리 이상이라는 보조적인 제안을 언급했습니다. 대신, 이들은 서로 이형적 상호작용에 참여하는 여러 별개의 세포 유형으로 구성된 복잡한 조직입니다. 우리는 종양 관련 간질을 형성하는 모집된 정상 세포를 수동적인 방관자가 아닌 종양 형성의 적극적인 참가자로 묘사했습니다. 따라서 이러한 간질 세포는 특정 특징 기능의 개발 및 발현에 기여합니다. 이후 10년 동안 이 개념은 확고해지고 확장되어 종양의 생물학은 더 이상 단순히 암세포의 특성을 열거하는 것만으로는 이해될 수 없으며 대신 종양 형성에 대한 “종양 미세환경”의 기여를 포괄해야 한다는 사실이 밝혀졌습니다.
이 출판물 이후 암 연구가 눈부시게 발전하는 과정에서 새로운 관찰 결과는 특징적인 능력의 원래 공식을 명확하게 하고 수정하는 데 도움이 되었습니다. 또한, 다른 관찰에서는 특징적인 특성에 대한 원래의 정교화에 필수적이지 않은 의문점이 제기되고 기계적 개념이 강조되었습니다. 이러한 발전에 동기를 부여받아 우리는 이제 원래의 특징을 다시 살펴보고 이 명단에 포함될 수 있는 새로운 특징을 고려하며 모집된 간질 세포가 종양 생물학에 기여하는 기능적 역할과 기여를 확장합니다.
특징적인 기능 – 개념적 발전
암의 6가지 특징(종양 성장과 전이성 전파를 가능하게 하는 독특하고 보완적인 능력)은 계속해서 암의 생물학을 이해하기 위한 견고한 기반을 제공합니다( 그림 1, 보충 정보 참조 )프리젠테이션용 그림의 다운로드 가능한 버전). 이 리뷰의 첫 번째 섹션에서 우리는 2000년 원본 프레젠테이션에 설명된 대로 각 특징의 본질을 요약한 다음, 기계적 토대를 이해하는 데 있어 지난 10년 동안 이루어진 개념적 진보에 대한 선택된 그림(이탤릭체로 표시된 하위 제목으로 구분)을 요약합니다. 후속 섹션에서 우리는 개념화의 범위를 넓히는 새로운 개발을 다루며, 6가지 특징 기능을 획득하는 데 중요한 두 가지 활성화 특성, 두 가지 새로운 새로운 특징 기능, 암 표현형에 중요한 종양 미세 환경의 구성 및 신호 상호 작용을 설명합니다. 그리고 마침내 이러한 개념을 치료에 적용하는 새로운 영역에 대해 논의합니다.
증식성 신호 유지
아마도 암세포의 가장 기본적인 특성은 만성 증식을 유지하는 능력과 관련이 있습니다. 정상 조직은 세포 성장 및 분열 주기에 진입하고 진행하도록 지시하는 성장 촉진 신호의 생성과 방출을 신중하게 제어하여 세포 수의 항상성을 보장하고 이에 따라 정상적인 조직 구조와 기능을 유지합니다. 암세포는 이러한 신호의 조절을 완화함으로써 자신의 운명의 주인이 됩니다. 활성화 신호는 일반적으로 세포내 티로신 키나제 도메인을 포함하는 세포 표면 수용체에 결합하는 성장 인자에 의해 대부분 전달됩니다. 후자는 세포 주기뿐만 아니라 세포 성장을 통한 진행을 조절하는 분기된 세포내 신호 전달 경로를 통해 신호를 방출합니다(즉, 세포 크기 증가); 종종 이러한 신호는 세포 생존 및 에너지 대사와 같은 다른 세포 생물학적 특성에 영향을 미칩니다.
놀랍게도 정상 조직 내에서 작동하는 증식 신호의 정확한 정체와 출처는 10년 전에는 제대로 이해되지 않았으며 일반적으로 여전히 그렇습니다. 더욱이, 우리는 이러한 유사 분열 신호의 방출을 제어하는 메커니즘에 대해 아직 상대적으로 거의 알지 못합니다. 부분적으로, 조직 내 세포 수와 위치를 제어하는 성장 인자 신호가 한 세포에서 이웃 세포로 시간적, 공간적으로 조절되는 방식으로 전달되는 것으로 생각된다는 사실로 인해 이러한 메커니즘을 이해하는 것이 복잡해집니다. 이러한 측분비 신호 전달은 실험적으로 접근하기 어렵습니다. 또한, 성장 인자의 생체 이용률은 세포주위 공간과 세포외 기질의 격리와 프로테아제, 설파타제, 대조적으로, 암세포의 분열촉진 신호전달은 더 잘 이해되고 있습니다. (레몬과 슐레진저, 2010,위치 외., 2010,하인즈와 맥도날드, 2009,페로나, 2006). 암세포는 다양한 대체 방법으로 증식 신호를 유지하는 능력을 획득할 수 있습니다. 즉, 성장 인자 리간드를 자체적으로 생성할 수 있으며, 이에 동족 수용체의 발현을 통해 반응하여 자가분비 증식 자극을 일으킬 수 있습니다. 대안적으로, 암세포는 지지하는 종양 관련 간질 내의 정상 세포를 자극하는 신호를 보낼 수 있으며, 이는 암세포에 다양한 성장 인자를 공급함으로써 상호 작용합니다. (쳉 외, 2008, Bhowmick 외, 2004). 수용체 신호전달은 또한 암세포 표면에 표시되는 수용체 단백질의 수준을 높여 이러한 세포가 제한된 양의 성장 인자 리간드에 과민반응하도록 함으로써 규제를 완화할 수 있습니다. 리간드 독립적 발사를 촉진하는 수용체 분자의 구조적 변화로 인해 동일한 결과가 발생할 수 있습니다.
성장 인자 독립성은 또한 이들 수용체의 하류에서 작동하는 신호 전달 경로 구성 요소의 구성적 활성화로부터 유래할 수 있으며, 이는 리간드 매개 수용체 활성화에 의해 이들 경로를 자극할 필요성을 제거합니다. 다수의 별개의 하류 신호 전달 경로가 리간드 자극 수용체로부터 방사된다는 점을 고려하면, 이러한 하류 경로 중 하나 또는 다른 것, 예를 들어 Ras 신호 변환기에 반응하는 신호의 활성화는 활성화된 수용체에 의해 전송된 조절 명령의 하위 집합만을 요약할 수 있습니다.
체세포 돌연변이는 추가적인 하류 경로를 활성화합니다
암세포 게놈에 대한 높은 처리량의 DNA 염기서열 분석을 통해 일반적으로 활성화된 성장 인자 수용체에 의해 유발되는 신호 회로의 구성적 활성화를 예측하는 특정 인간 종양의 체세포 돌연변이가 밝혀졌습니다. 따라서 우리는 이제 인간 흑색종의 약 40%가 B-Raf 단백질의 구조에 영향을 미치는 활성화 돌연변이를 포함하고 있으며, 그 결과 Raf를 통해 미토겐 활성화 단백질(MAP)-키나제 경로로의 구성적 신호 전달이 이루어진다는 것을 알고 있습니다. (데이비스와 사무엘스, 2010). 유사하게, 포스포이노시티드 3-키나제(PI3-키나제) 이소형의 촉매 서브유닛의 돌연변이가 다양한 종양 유형에서 검출되고 있으며, 이는 주요 Akt/PKB 신호 변환기를 포함하여 PI3-키나제 신호 전달 회로를 과도하게 활성화하는 역할을 합니다. (장과 리우, 2009, 위안과 캔틀리, 2008). 성장인자 수용체에서 방출되는 다중 경로 사이의 누화의 기능적 영향과 마찬가지로 상류(수용체) 신호와 하류(변환기) 신호를 활성화하는 종양 세포의 이점은 여전히 불분명합니다.
증식성 신호를 약화시키는 부정적인 피드백 메커니즘의 중단
최근 결과는 일반적으로 다양한 유형의 신호 전달을 약화시켜 세포내 회로를 통과하는 신호 흐름의 항상성 조절을 보장하는 네거티브 피드백 루프의 중요성을 강조했습니다. (워츠와 딕시트, 2010, 카브리타와 크리스토포리, 2008,아미트 외., 2007,모세슨 외, 2008). 이러한 피드백 메커니즘의 결함은 증식 신호를 향상시킬 수 있습니다. 이러한 유형의 조절의 원형은 Ras 종양단백질과 관련이 있습니다. Ras의 발암 효과는 신호 능력의 과잉 활성화로 인해 발생하지 않습니다. 대신, ras 유전자에 영향을 미치는 발암성 돌연변이는 Ras GTPase 활성을 손상시킵니다. 이는 일반적으로 활성 신호 전송이 일시적임을 보장하는 본질적인 음성 피드백 메커니즘으로 작동합니다.
유사한 네거티브 피드백 메커니즘은 증식 신호 회로 내의 여러 노드에서 작동합니다. 눈에 띄는 예는 PTEN 포스파타제와 관련이 있는데, 이는 해당 생성물인 포스파티딜이노시톨(3,4,5) 삼인산(PIP 3 )을 분해하여 PI3-키나제에 대응합니다. PTEN의 기능 상실 돌연변이는 PI3K 신호를 증폭시키고 다양한 암 실험 모델에서 종양 형성을 촉진합니다. 인간 종양에서는 PTEN 발현이 프로모터 메틸화에 의해 손실되는 경우가 많습니다. (장과 리우, 2009, 위안과 캔틀리, 2008).
또 다른 예는 PI3K 경로의 상류와 하류 모두에 존재하는 세포 성장 및 대사의 조정자인 mTOR 키나제와 관련이 있습니다. 일부 암세포의 회로에서 mTOR 활성화는 음성 피드백을 통해 PI3K 신호 전달을 억제합니다. 따라서 mTOR이 그러한 암세포에서 약리학적으로 억제되면(예: 라파마이신 약물에 의해) 음성 피드백의 관련 손실로 인해 PI3K 및 그 효과기 Akt/PKB의 활성이 증가하여 mTOR 억제의 항증식 효과가 둔화됩니다. (수다르사남과 존슨, 2010년, 오라일리 외, 2006). 이 신호 경로와 다른 신호 전달 경로에서 손상된 음성 피드백 루프는 인간 암세포 사이에 널리 퍼져 있으며 이러한 세포가 증식 독립성을 달성할 수 있는 중요한 수단으로 작용할 가능성이 높습니다. 더욱이, 이러한 자가 감쇠 신호 전달의 중단은 분열 촉진 신호 전달을 표적으로 하는 약물에 대한 적응성 저항의 발달에 기여할 수 있습니다.
과도한 증식 신호는 세포 노화를 유발할 수 있습니다
종양유전자 작용에 대한 초기 연구는 그러한 유전자의 발현이 계속 증가하고 단백질 생성물에서 나타나는 신호가 그에 따라 암세포 증식을 증가시켜 종양 성장을 초래할 것이라는 개념을 장려했습니다. 보다 최근의 연구에서는 RAS, MYC 및 RAF와 같은 종양단백질에 의한 신호 전달의 과도하게 증가가 세포의 반작용 반응, 특히 세포 노화 및/또는 세포사멸의 유도를 유발할 수 있다는 점에서 이러한 개념을 약화시켰습니다. (콜라도와 세라노, 2010, 에반과 다다 디 파가냐, 2009, 로우 외., 2004). 예를 들어, 높은 수준의 Ras 종양단백질을 발현하는 배양된 세포는 노화라고 불리는 비증식성이지만 생존 가능한 상태로 들어갈 수 있습니다. 대조적으로, 이 단백질의 낮은 수준을 발현하는 세포는 노화를 피하고 증식할 수 있습니다.
세포질 확대, 증식 표지의 부재, 노화 유발 베타-갈락토시다제 효소의 발현 등 노화의 형태학적 특징을 지닌 세포가 특정 종양 유전자를 과발현하도록 조작된 쥐의 조직에 풍부하게 존재합니다. (콜라도와 세라노, 2010,에반과 다다 디 파가냐, 2009) 인간 흑색종의 일부 사례에서 널리 퍼져 있습니다(무이와 피퍼, 2006). 이러한 표면적으로 역설적인 반응은 과도한 수준의 특정 유형의 신호 전달을 경험하는 세포를 제거하도록 설계된 본질적인 세포 방어 메커니즘을 반영하는 것으로 보입니다. 따라서, 암세포에서 발암성 신호 전달의 상대적 강도는 최대 분열 촉진 자극과 이러한 항증식 방어의 회피 사이의 절충안을 나타낼 수 있습니다. 또는 일부 암세포는 노화 또는 세포사멸 유도 회로를 비활성화하여 높은 수준의 발암성 신호 전달에 적응할 수 있습니다.
성장억제제 회피
암세포는 긍정적으로 작용하는 성장 자극 신호를 유도하고 유지하는 특징적인 능력 외에도 세포 증식을 부정적으로 조절하는 강력한 프로그램을 피해야 합니다. 이들 프로그램 중 다수는 종양 억제 유전자의 작용에 의존합니다. 세포 성장과 증식을 제한하기 위해 다양한 방식으로 작동하는 수십 가지의 종양 억제 인자가 하나 또는 다른 형태의 동물 또는 인간 암에서 특징적인 불활성화를 통해 발견되었습니다. 이들 유전자 중 다수는 생쥐의 기능 획득 또는 상실 실험을 통해 진정한 종양 억제 인자로 검증되었습니다. 두 가지 원형 종양 억제인자는 RB(망막모세포종 관련) 및 TP53 단백질을 암호화합니다. 이들은 세포의 증식 또는 노화 및 세포사멸 프로그램 활성화 결정을 통제하는 두 가지 주요 보완적인 세포 조절 회로 내에서 중앙 제어 노드로 작동합니다.
RB 단백질은 다양한 세포외 및 세포내 소스의 신호를 통합하고 그에 대한 응답으로 세포가 성장과 분열 주기를 진행해야 하는지 여부를 결정합니다. (버크하트와 세이지, 2008, Deshpande 외, 2005, 셰르와 맥코믹, 2002). 따라서 RB 경로 기능에 결함이 있는 암세포는 지속적인 세포 증식을 허용하는 세포주기 진행의 중요한 문지기 역할을 놓치게 됩니다. RB가 주로 세포 외부에서 발생하는 성장 억제 신호를 전달하는 반면, TP53은 게놈 손상 정도가 과도한 경우 또는 뉴클레오티드 풀 수준이 과도한 경우 세포의 세포내 운영 체제 내에서 기능하는 스트레스 및 이상 센서로부터 입력을 받습니다. 성장 촉진 신호, 포도당 또는 산소 공급이 최적이 아닌 경우, TP53은 이러한 조건이 정상화될 때까지 추가 세포 주기 진행을 중단할 수 있습니다. 대안적으로, 그러한 세포 하위 시스템에 대한 압도적이거나 회복 불가능한 손상을 나타내는 경보 신호에 직면하여 TP53은 세포 사멸을 유발할 수 있습니다. 특히, 두 가지 표준적인 증식 억제 인자인 TP53과 RB는 세포 증식을 조절하는 데 탁월한 중요성을 갖고 있지만, 다양한 증거에 따르면 각각은 기능적 중복을 위해 연결된 더 큰 네트워크의 일부로 작동하는 것으로 나타났습니다. 예를 들어, 기능성 Rb 유전자가 결여된 개별 세포로 몸 전체에 거주하는 키메라 쥐는 RB 기능의 상실로 인해 이들 세포와 직계 후손에서 세포 분열 주기가 지속적으로 시작될 것이라는 기대에도 불구하고 놀랍게도 증식 이상이 없습니다. 결과적으로 발생하는 Rb null 세포 클러스터 중 일부는 신생물로 진행되어야 합니다. 대신 Rb는 이러한 키메라 쥐의 null 세포는 신체 전반에 걸쳐 상대적으로 정상적인 조직 형태 형성에 참여하는 것으로 밝혀졌습니다. 관찰된 유일한 신생물은 말년에 뇌하수체 종양이 발생한 것이었습니다(리핀스키와 잭스, 1999). 유사하게, TP53이 없는 쥐는 정상적으로 발달하고 대체로 적절한 세포와 조직의 항상성을 보여주다가 나중에 백혈병과 육종의 형태로 다시 이상을 일으킨다. (게브라니우스와 돈하워, 1998). 두 사례 모두 이러한 주요 증식 억제 인자가 결여된 세포의 부적절한 복제를 제한하는 역할을 하는 중복 작용 메커니즘의 작동을 반영해야 합니다.
접촉 억제 및 회피 메커니즘
40년 간의 연구를 통해 2차원 배양에서 증식된 정상 세포의 밀집된 집단에 의해 형성된 세포 간 접촉이 추가 세포 증식을 억제하여 융합성 세포 단층을 생성한다는 것이 입증되었습니다. 중요한 것은 이러한 “접촉 억제”가 배양된 다양한 유형의 암세포에서 폐지된다는 점입니다. 이는 접촉 억제가 정상적인 조직 항상성을 보장하기 위해 생체 내에서 작동하는 메커니즘의 시험관 내 대리자이며 종양 형성 과정에서 폐지되는 메커니즘임을 시사합니다. 최근까지 이러한 성장 조절 방식의 기계적 기초는 여전히 불분명했습니다. 그러나 이제 접촉 억제 메커니즘이 나타나기 시작했습니다.
한 가지 메커니즘은 NF2 유전자의 산물과 관련이 있는데, 그 손실은 인간 신경섬유종증의 형태를 유발하기 때문에 오랫동안 종양 억제 인자로 연루되어 왔습니다. 세포질 NF2 유전자 산물인 Merlin은 세포 표면 접착 분자(예: E-cadherin)를 막횡단 수용체 티로신 키나제(예: EGF 수용체)에 결합시켜 접촉 억제를 조정합니다. 그렇게 함으로써 Merlin은 카드헤린 매개 세포 간 부착의 접착성을 강화합니다. 또한, 성장 인자 수용체를 격리함으로써 Merlin은 분열 촉진 신호를 효율적으로 방출하는 능력을 제한합니다. (커토 등, 2007,오카다 외, 2005).
접촉 억제의 두 번째 메커니즘은 상피 구조를 구성하고 조직 무결성을 유지하는 데 도움이 되는 LKB1 상피 극성 단백질과 관련이 있습니다. 예를 들어, LKB1은 강력한 Myc 종양유전자가 조직화되고 정지된 상피 구조에서 상향조절될 때 이의 유사분열 효과를 무시할 수 있습니다. 대조적으로, LKB1 발현이 억제되면 상피 완전성이 불안정해지고 상피 세포는 Myc 유발 형질전환에 취약해집니다. (파르타넨 외, 2009,헤젤과 바디시, 2008). LKB1은 또한 특정 인간 악성 종양에서 상실되는 종양 억제 유전자로 확인되었습니다. (쇼, 2009), 아마도 부적절한 증식을 억제하는 정상적인 기능을 반영하는 것 같습니다. 접촉 매개 성장 억제의 이 두 가지 메커니즘이 인간 암에서 얼마나 자주 손상되는지는 아직 알 수 없습니다. 의심할 바 없이 아직 다른 접촉으로 인한 증식 장벽은 아직 발견되지 않았습니다. 세포가 구조적으로 복잡한 조직을 구성하고 유지할 수 있도록 하는 이와 같은 메커니즘은 부적절한 증식 신호를 억제하고 균형을 맞추는 중요한 수단을 나타냅니다.
TGF-β 경로의 손상은 악성종양을 촉진합니다
TGF-β는 항증식 효과로 가장 잘 알려져 있으며 암세포가 이러한 효과를 회피하는 것은 신호 회로를 단순히 차단하는 것보다 훨씬 더 정교한 것으로 알려져 있습니다. (이쿠시마와 미야조노, 2010년,마사게, 2008,비에리와 모세스, 2006). 많은 말기 종양에서 TGF-β 신호 전달은 세포 증식을 억제하는 방향에서 벗어나 대신 상피에서 중간엽으로의 전환(EMT)이라고 불리는 세포 프로그램을 활성화하는 것으로 밝혀졌습니다. 등급 악성종양은 아래에서 더 자세히 논의됩니다.
세포 사멸 저항
세포사멸에 의한 프로그램화된 세포 사멸이 암 발병에 대한 자연적 장벽 역할을 한다는 개념은 지난 20년 동안 수행된 기능적 연구를 통해 확립되었습니다. (아담스와 코리, 2007,로우 외., 2004:에반과 리틀우드, 1998). 세포사멸 프로그램을 관장하는 신호 회로의 해명은 종양 형성 과정 동안 또는 항암 치료의 결과로 암세포가 경험하는 다양한 생리적 스트레스에 반응하여 세포사멸이 어떻게 유발되는지 밝혀졌습니다. 세포사멸을 유발하는 스트레스 중에서 주목할 만한 것은 앞서 언급한 바와 같이 종양유전자 신호의 증가된 수준으로 인한 신호 불균형과 과다증식과 관련된 DNA 손상입니다. 그러나 다른 연구에서는 높은 등급의 악성 종양 상태와 치료에 대한 저항성 상태로 성공적으로 진행된 종양에서 세포사멸이 어떻게 약화되는지 밝혀냈습니다. (아담스와 코리, 2007,로우 외., 2004).
세포사멸 기계는 상류 조절자와 하류 효과기 구성 요소로 구성됩니다. (아담스와 코리, 2007). 조절자는 차례로 두 개의 주요 회로로 나누어집니다. 하나는 세포외 사멸 유도 신호(예를 들어 Fas 리간드/Fas 수용체를 포함하는 외인성 세포사멸 프로그램)를 수신하고 처리하는 회로이고, 다른 하나는 다양한 신호를 감지하고 통합하는 회로입니다. 세포내 기원(내재적 프로그램). 각각은 일반적으로 잠재되어 있는 프로테아제(각각 카스파제 8과 9)의 활성화로 최고조에 이르며, 이는 세포사멸의 실행 단계를 담당하는 이펙터 카스파제를 포함하는 일련의 단백질 분해를 시작합니다. 여기서 세포는 점진적으로 분해된 다음 소비됩니다. 그 이웃과 전문적인 식세포에 의해. 현재, 내인성 세포사멸 프로그램은 암 발병에 대한 장벽으로 더 광범위하게 관련되어 있습니다.
조절자와 작동자 사이에 신호를 전달하는 “세포사멸 유발 요인”은 Bcl-2 조절 단백질 계열의 세포사멸 촉진 및 항아포토시스 구성원의 균형을 맞춰 제어됩니다. (아담스와 코리, 2007). 원형인 Bcl-2는 가장 가까운 친척(Bcl-x L , Bcl-w, Mcl-1, A1)과 함께 세포 사멸 억제제이며, 두 가지 proapoptotic 유발 단백질(Bax 및 Bak); 후자는 미토콘드리아 외막에 내장되어 있습니다. 항아포토시스 친척에 의한 억제가 완화되면 Bax와 Bak는 외부 미토콘드리아 막의 완전성을 파괴하여 아포토시스 촉진 신호 단백질(가장 중요한 것은 시토크롬 c) 의 방출을 유발합니다. 방출된 시토크롬 c는 차례로 세포사멸 프로그램과 관련된 다중 세포 변화를 유도하기 위해 단백질 분해 활동을 통해 작용하는 카스파제의 폭포를 활성화합니다. Bax와 Bak는 BH3 모티프라고 불리는 단백질-단백질 상호 작용 도메인을 다양한 물리적 상호 작용을 중재하는 항아포토시스 Bcl-2 유사 단백질과 공유합니다. 각각 단일 BH3 모티프를 포함하는 관련 단백질 서브패밀리의 활동은 다양한 세포 이상 센서와 결합됩니다. 이러한 “BH3-only” 단백질은 세포사멸 방지 Bcl-2 단백질을 방해하거나 이 계열의 세포사멸 촉진 구성원을 직접 자극함으로써 작용합니다. (아담스와 코리, 2007,윌리스와 아담스, 2005).
세포사멸을 유발하는 세포 조건은 완전히 열거되지 않았지만 종양 발달에 중요한 역할을 하는 몇 가지 이상 센서가 확인되었습니다. (아담스와 코리, 2007,로우 외., 2004). 가장 주목할만한 것은 TP53 종양 억제자를 통해 기능하는 DNA 손상 센서입니다. (준틸라와 에반, 2009년); TP53은 Noxa 및 Puma BH3 전용 단백질의 발현을 상향 조절하여 상당한 수준의 DNA 파손 및 기타 염색체 이상에 반응하여 세포사멸을 유도합니다. 대안적으로, 불충분한 생존 인자 신호 전달(예를 들어 림프구의 인터루킨-3 또는 상피 세포의 인슐린 유사 성장 인자 1/2[Igf1/2]의 부적절한 수준)은 Bim이라는 BH3 전용 단백질을 통해 세포사멸을 유발할 수 있습니다. 세포 사멸을 초래하는 또 다른 조건은 항세포사멸 인자에 의해 균형을 이루지 않는 한 세포사멸을 유발하는(부분적으로 Bim 및 기타 BH3 전용 단백질을 통해) Myc와 같은 특정 종양 단백질에 의한 과잉 활성 신호 전달과 관련이 있습니다. (준틸라와 에반, 2009년,로우 외., 2004).
종양 세포는 세포사멸을 제한하거나 회피하기 위해 다양한 전략을 발전시킵니다. 가장 흔한 것은 TP53 종양 억제 기능의 상실로, 이는 세포사멸 유도 회로에서 이 중요한 손상 센서를 제거합니다. 대안적으로, 종양은 항아폽토시스 조절인자(Bcl-2, Bcl- xL ) 또는 생존 신호(Igf1/2) 의 발현을 증가시키거나, 아폽토시스 촉진 인자(Bax, Bim, Puma)를 하향 조절하거나 단락을 통해 유사한 목적을 달성할 수 있습니다. 외부 리간드에 의한 사망 경로. 세포사멸을 방지하는 메커니즘의 다양성은 아마도 암세포 집단이 악성 상태로 진화하는 동안 직면하는 세포사멸 유도 신호의 다양성을 반영하는 것 같습니다.
세포사멸 기계와 프로그램의 구조, 그리고 암세포가 그 작용을 회피하기 위해 사용하는 전략은 지난 10년 초에 널리 인식되었습니다. 그 이후로 가장 주목할 만한 개념적 발전은 암에 대한 장벽으로서 “프로그램화된 세포 사멸”의 범위를 넓히는 다른 형태의 세포 사멸과 관련이 있었습니다.
자가포식은 종양 세포의 생존과 죽음을 모두 중재합니다
자가포식은 세포사멸과 마찬가지로 일반적으로 세포의 낮은 기초 수준에서 작동하지만 특정 세포 스트레스 상태에서 강력하게 유도될 수 있는 중요한 세포 생리적 반응을 나타내며, 그 중 가장 명백한 것은 영양 결핍입니다. (레빈과 크로머, 2008,미즈시마, 2007). 자가포식 프로그램은 세포가 리보솜 및 미토콘드리아와 같은 세포 소기관을 분해할 수 있게 하여 생성된 분해 산물이 재활용되어 생합성 및 에너지 대사에 사용될 수 있도록 합니다. 이 프로그램의 일부로, 자가포식소체라고 불리는 세포내 소포는 세포내 소기관을 둘러싸서 분해가 일어나는 리소좀과 융합합니다. 이러한 방식으로 많은 암세포가 겪는 스트레스와 영양이 제한된 환경에서 생존을 지원하는 저분자량 대사산물이 생성됩니다.
세포사멸과 마찬가지로, 자가포식 기구는 조절 및 효과기 구성 요소를 모두 가지고 있습니다. (레빈과 크로머, 2008,미즈시마, 2007). 후자 중에는 자가포식소체 형성과 리소좀으로의 전달을 중재하는 단백질이 있습니다. 주목할 만한 최근 연구에서는 자가포식, 세포사멸, 세포 항상성을 관장하는 조절 회로 사이의 교차점이 밝혀졌습니다. 예를 들어, 생존 신호에 의해 자극되어 세포사멸을 차단하는 PI3-키나제, AKT 및 mTOR 키나제와 관련된 신호 전달 경로는 유사하게 자가포식을 억제합니다. 생존 신호가 충분하지 않으면 PI3K 신호 전달 경로가 하향 조절되어 자가포식 및/또는 세포사멸이 유도될 수 있습니다. (레빈과 크로머, 2008,신하와 레빈, 2008,매튜 등, 2007).
이 두 프로그램 사이의 또 다른 상호 연결은 Beclin-1 단백질에 있는데, 이는 유전학 연구를 통해 자가포식 유도에 필요한 것으로 나타났습니다. (레빈과 크로머, 2008,신하와 레빈, 2008,미즈시마, 2007). Beclin-1은 세포사멸 조절 단백질의 BH3 전용 서브패밀리의 구성원이며, Beclin-1의 BH3 도메인은 Bcl-2/Bcl-x L 단백질과 결합할 수 있게 해줍니다. 스트레스 센서 결합 BH3 단백질은 Beclin-1을 Bcl-2/Bcl-x L 과의 결합에서 대체하여 해방된 Beclin-1이 자가포식을 촉발할 수 있도록 하며, 이는 세포사멸 촉진 Bax 및 Bak을 방출하여 세포사멸을 유발할 수 있는 것과 같습니다. 따라서 스트레스를 전달하는 BH3 단백질(예: Bid, Bad, Puma 등)은 세포의 생리적 상태에 따라 세포사멸 및/또는 자가포식을 유도할 수 있습니다.
Beclin-1 유전자나 자가포식 기구의 다른 특정 구성요소의 비활성화된 대립유전자를 보유한 쥐는 암에 대한 감수성이 증가한 것으로 나타났습니다. (화이트와 디파올라, 2009:레빈과 크로머, 2008). 이러한 결과는 자가포식의 유도가 세포사멸과 독립적으로 또는 동시에 작용할 수 있는 종양 형성에 대한 장벽 역할을 할 수 있음을 시사합니다. 따라서, 자가포식은 종양 발달 과정에서 우회해야 할 또 다른 장벽을 나타내는 것으로 보입니다. (화이트와 디파올라, 2009).
아마도 역설적이게도 영양 결핍, 방사선 요법 및 특정 세포 독성 약물은 암세포에 대한 세포 보호 효과가 있는 것으로 보이는 자가포식의 수준을 높여 이러한 스트레스 유발 상황의 살해 작용을 강조하기보다는 손상시킬 수 있습니다. (화이트와 디파올라, 2009,아펠 외., 2009,아마라바디와 톰슨, 2007,매튜 등, 2007). 더욱이, 심각한 스트레스를 받은 암세포는 자가포식을 통해 가역적인 휴면 상태로 축소되는 것으로 나타났습니다. (화이트와 디파올라, 2009,루 외., 2008). 이러한 생존 반응은 강력한 항암제로 치료한 후 일부 말기 종양의 지속성과 궁극적인 재성장을 가능하게 할 수 있습니다. 따라서, 종양 형성의 초기 단계에서는 종양을 억제하고 나중에는 종양을 촉진할 수 있는 TGF-β 신호 전달과 유사하게, 자가포식은 종양 세포에 상충되는 효과를 가져서 종양 진행에 영향을 미치는 것으로 보입니다. (아펠 외., 2009,화이트와 디파올라, 2009). 미래 연구의 중요한 의제는 자가포식을 통해 암세포가 생존하거나 죽게 만드는 시기와 방법을 결정하는 유전적 및 세포 생리학적 조건을 명확히 하는 것입니다.
괴사는 전염증성 및 종양 촉진 가능성이 있습니다.
죽어가는 세포가 거의 보이지 않는 시체로 수축되어 곧 이웃에 의해 소비되는 세포사멸과 달리, 괴사 세포는 부풀어 오르고 폭발하여 내용물을 국소 조직 미세 환경으로 방출합니다. 괴사는 역사적으로 유기체의 죽음과 매우 유사하게 시스템 전체의 고갈과 붕괴의 한 형태로 간주되었지만 개념적 지형은 변화하고 있습니다. 괴사에 의한 세포 죽음은 무작위적이고 방향이 없는 과정이 아닌 어떤 상황에서는 확실히 유전적 통제하에 있습니다. (Galluzzi와 Kroemer, 2008,종과 톰슨, 2006).
아마도 더 중요한 것은, 괴사성 세포 사멸은 그렇지 않은 세포사멸 및 자가포식과는 대조적으로 염증성 신호를 주변 조직 미세환경으로 방출한다는 것입니다. 결과적으로, 괴사 세포는 면역 체계의 염증 세포를 모집할 수 있습니다. (그리벤니코프 외, 2010,화이트 외, 2010,Galluzzi와 Kroemer, 2008)의 전용 기능은 조직 손상 정도를 조사하고 관련 괴사 잔해를 제거하는 것입니다. 그러나 신생물과 관련하여 여러 가지 증거는 면역 염증 세포가 혈관 신생, 암 세포 증식 및 침습성을 촉진할 수 있다는 점을 고려하면 면역 염증 세포가 적극적으로 종양을 촉진할 수 있음을 나타냅니다(아래 참조). 또한, 괴사성 세포는 IL-1α와 같은 생리활성 조절 인자를 방출할 수 있는데, 이는 인접한 생존 세포의 증식을 직접 자극하여 종양성 진행을 촉진할 가능성이 있습니다. (그리벤니코프 외, 2010). 결과적으로, 괴사성 세포 사멸은 암과 관련된 과잉 증식을 상쇄하는 데 도움이 되는 것처럼 보이지만 궁극적으로는 좋은 것보다 더 많은 피해를 줄 수 있습니다. 따라서, 초기 신생물과 잠재적으로 침윤성 및 전이성 종양은 괴사성 세포 사멸을 어느 정도 용인함으로써 이점을 얻을 수 있으며, 이는 이러한 성장 내에서 살아남은 세포에 성장 자극 인자를 가져오는 종양 촉진 염증 세포를 모집하기 위한 것입니다.
복제 불멸성 활성화
2000년까지 암세포가 육안으로 보이는 종양을 생성하려면 무제한의 복제 잠재력이 필요하다는 것이 널리 받아들여졌습니다. 이 능력은 제한된 수의 연속적인 세포 성장 및 분열 주기만 통과할 수 있는 신체의 대부분의 정상적인 세포 계통의 세포 행동과 현저한 대조를 이룹니다. 이러한 제한은 증식에 대한 두 가지 뚜렷한 장벽, 즉 일반적으로 비증식이지만 생존 가능한 상태로 돌이킬 수 없는 진입인 노화와 세포 사멸을 수반하는 위기와 관련이 있습니다. 따라서, 세포가 배양에서 증식될 때, 세포 분열의 반복된 주기는 먼저 노화를 유도한 다음, 이 장벽을 우회하는 데 성공한 세포의 경우 집단의 대부분의 세포가 죽는 위기 단계로 이어집니다. 드문 경우지만, 세포는 위기에 처한 개체군에서 나타나며 무한한 복제 잠재력을 나타냅니다. 이러한 전환을 불멸화(immortalization)라고 부르는데, 이는 대부분의 확립된 세포주가 노화나 위기의 증거 없이 배양에서 증식하는 능력으로 인해 보유하는 특성입니다.
여러 증거에 따르면 염색체 말단을 보호하는 텔로미어가 무제한 증식 능력에 중심적으로 관여하고 있음이 밝혀졌습니다. (블라스코, 2005,셰이와 라이트, 2000). 여러 개의 직렬 헥사뉴클레오티드 반복으로 구성된 텔로미어는 배양 중에 증식된 불멸화되지 않은 세포에서 점진적으로 짧아지며 결국 염색체 DNA의 끝을 종단 간 융합으로부터 보호하는 능력을 잃습니다. 이러한 융합은 불안정한 이심성 염색체를 생성하며, 그 분해로 인해 세포 생존 능력을 위협하는 핵형이 뒤섞이게 됩니다. 따라서 세포 내 텔로미어 DNA의 길이는 텔로미어가 대부분 침식되어 결과적으로 보호 기능을 상실하여 위기에 빠지기 전에 그 자손이 얼마나 많은 연속적인 세포 세대를 통과할 수 있는지를 나타냅니다.
텔로미어 DNA의 말단에 텔로미어 반복 세그먼트를 추가하는 특수 DNA 중합효소인 텔로머라제는 비불멸화 세포에는 거의 없지만 인간 암세포를 포함하여 자연 불멸화 세포의 대다수(약 90%)에서는 기능적으로 중요한 수준으로 발현됩니다. 텔로미어 DNA를 확장함으로써 텔로머라제는 텔로미어가 없을 경우 발생할 수 있는 진행성 텔로미어 침식에 대응할 수 있습니다. 자발적으로 불멸화된 세포 또는 효소를 발현하도록 조작된 세포의 맥락에서 텔로머라제 활성의 존재는 노화 및 위기/세포사멸 유도에 대한 저항성과 상관관계가 있습니다. 반대로, 텔로머라제 활성이 억제되면 텔로미어가 짧아지고 이러한 증식 장벽 중 하나가 활성화됩니다.
증식에 대한 두 가지 장벽인 노화와 위기/세포사멸은 우리 세포에 내장되어 있는 중요한 항암 방어 수단으로 합리화되어 전종양 세포와 솔직히 신생물 세포 클론의 성장을 방해하기 위해 배치되었습니다. 이러한 생각에 따르면, 대부분의 초기 신생물은 복제 배가의 부여를 소진하고 이러한 장벽 중 하나 또는 다른 것에 의해 경로가 중단됩니다. 종양을 형성하는 희귀 변이 세포의 궁극적인 불멸화는 노화 또는 세포사멸을 유발하는 것을 피하기에 충분한 길이로 텔로미어 DNA를 유지하는 능력에 기인합니다. 가장 일반적으로는 텔로머라제의 발현을 상향 조절하거나 덜 자주 대체 재조합을 통해 달성됩니다. 텔로미어 유지 메커니즘을 기반으로 합니다. 따라서,
복제 노화 재평가
텔로미어 유지가 신생물 상태에 중요한 조건으로 점점 더 입증되고 있는 반면, 일반적인 장벽으로서의 복제 유발 노화의 개념은 개선과 재구성이 필요합니다. (마우스 세포와 인간 세포의 텔로미어 구조 및 기능의 차이로 인해 복제 노화에서 텔로미어와 텔로머라제의 역할에 대한 조사가 복잡해졌습니다.) 최근 실험에 따르면 특정 배양 세포의 노화 유도가 지연되거나 제거될 수 있음이 밝혀졌습니다. 개선된 세포 배양 조건은 최근에 이식된 일차 세포가 위기 지점까지 배양에서 방해받지 않고 증식할 수 있으며, 결정적으로 단축된 텔로미어에 의해 유발되는 세포사멸 유도와 관련될 수 있음을 시사합니다. (Ince 등, 2007,파소스 외, 2007,장 외., 2004,셰르(Sherr)와 드피뉴(DePinho), 2000년). 대조적으로, 텔로머라제가 결여되도록 조작된 쥐를 대상으로 한 실험에서는 결과적으로 단축된 텔로미어가 전악성 세포를 노화 상태로 전환시켜 (세포사멸과 함께) 유전적으로 특정 형태의 암이 발생하도록 운명지어진 쥐의 종양 발생을 약화시키는 데 기여할 수 있음을 나타냅니다. (아르탄디와 드피뉴, 2010). 심하게 침식된 텔로미어를 가진 이러한 텔로머라제 기능이 없는 쥐는 노화와 세포사멸에 대한 증거를 포함하는 다기관 기능 장애와 이상을 나타내며, 아마도 세포 배양에서 관찰되는 노화와 세포사멸과 유사할 것입니다. (아르탄디와 드피뉴, 2010,펠더와 그라이더, 2007).
주목할 만한 점은 앞서 논의한 바와 같이 과도하거나 불균형한 종양 유전자 신호 전달에 의해 유발된 형태학적으로 유사한 형태의 세포 노화가 이제 신생물에 대한 보호 메커니즘으로 잘 문서화되어 있다는 것입니다. 이러한 형태의 노화와 텔로머라제 및 텔로미어의 가능한 상호 연결은 아직 확인되지 않았습니다. 따라서 세포 노화는 개념적으로 높은 수준의 발암성 신호 전달 및 임계 미만의 텔로미어 단축을 비롯한 다양한 증식 관련 이상에 의해 유발될 수 있는 종양 확장에 대한 보호 장벽으로 떠오르고 있습니다.
텔로머라제의 활성화 지연은 종양성 진행을 제한하고 촉진할 수 있습니다.
이제 초기 암세포의 클론은 상당한 수준의 텔로머라제를 발현할 수 없기 때문에 다단계 종양 진행 과정에서 상대적으로 초기에 텔로미어 손실로 인한 위기를 경험한다는 증거가 있습니다. 따라서 광범위하게 침식된 텔로미어는 형광 in situ 하이브리드화(FISH)를 사용하여 전암성 성장에서 기록되었으며, 이는 또한 텔로미어 실패와 위기를 알리는 종단간 염색체 융합을 밝혀냈습니다. (카와이 외, 2007,헨젤 등, 2006). 이러한 결과는 또한 그러한 세포가 완전히 정상적인 기원 세포에서 진화하는 동안 상당한 수의 연속적인 텔로미어 단축 세포 분열을 거쳤음을 시사합니다. 따라서 일부 인간 신생물의 발달은 거시적이고 솔직한 신생물 성장이 되기 훨씬 전에 텔로미어에 의해 유발된 위기에 의해 중단될 수 있습니다.
대조적으로, 게놈 무결성에 대한 TP53 매개 감시가 없으면 다른 초기 신생물이 초기 텔로미어 침식 및 그에 따른 염색체 파손-융합-교량(BFB) 주기에서 살아남을 수 있습니다. 염색체 부분의 결실 및 증폭을 포함하여 이러한 BFB 주기로 인한 게놈 변경은 분명히 게놈의 돌연변이성을 증가시켜 돌연변이 종양 유전자 및 종양 억제 유전자의 획득을 가속화하는 역할을 합니다. 손상된 텔로미어 기능이 실제로 종양 진행을 촉진할 수 있다는 사실은 p53과 텔로머라제 기능이 모두 결여된 돌연변이 쥐에 대한 연구에서 나왔습니다. (아르탄디와 드피뉴, 2010,아르탄디와 드피뉴, 2000). 이 두 가지 결함이 협력하여 인간 종양 형성을 향상시킬 수 있다는 제안은 아직 직접적으로 문서화되지 않았습니다.
악성 진행을 촉진하는데 있어서 일시적인 텔로미어 결핍의 중요성에 대한 정황적 뒷받침은 또한 인간 유방의 전악성 병변과 악성 병변에 대한 비교 분석에서 나왔습니다. (레이노 외., 2010,친 외., 2004). 전악성 병변은 상당한 수준의 텔로머라제를 발현하지 않았으며 텔로미어 단축 및 비클론성 염색체 이상으로 표시되었습니다. 대조적으로, 명백한 암종은 더 긴 텔로미어의 재구성 및 텔로미어 실패 이후에 텔로머라제 활성이 획득되기 전에 획득된 것으로 보이는 비정상적인 핵형의 고정(클론 파생물을 통해)과 일치하게 텔로머라제 발현을 나타냈습니다. 이러한 방식으로 묘사되면 텔로머라제 기능의 지연된 획득은 종양 촉진 돌연변이를 생성하는 역할을 하는 반면, 후속 활성화는 돌연변이 게놈을 안정화하고 암세포가 임상적으로 명백한 종양을 생성하기 위해 필요한 무제한 복제 능력을 부여합니다.
텔로머라제의 새로운 기능
텔로머라제는 텔로미어 DNA를 늘리고 유지하는 능력 때문에 발견되었으며, 거의 모든 텔로머라제 연구는 그 기능이 이 중요한 기능에 국한되어 있다는 개념에 상정되어 왔습니다. 그러나 최근 몇 년 동안 텔로머라제가 세포 증식과 관련이 있지만 텔로미어 유지와는 관련이 없는 기능을 발휘한다는 것이 명백해졌습니다. 텔로머라제, 특히 텔로머라제의 단백질 서브유닛 TERT의 비정규적인 역할은 생쥐와 배양 세포의 기능적 연구를 통해 밝혀졌습니다. 어떤 경우에는 텔로머라제 효소 활성이 제거된 조건에서 새로운 기능이 입증되었습니다. (콩과 셰이, 2008). 점점 늘어나고 있는 TERT/텔로머라제의 텔로미어 독립적 기능 목록 중에는 β-카테닌/LEF 전사 인자 복합체의 보조 인자 역할을 함으로써 Wnt 경로에 의한 신호 전달을 증폭시키는 TERT의 능력이 있습니다. (박 외, 2009). 다른 텔로미어 독립적 효과에는 세포 증식 및/또는 세포사멸에 대한 저항성의 입증 가능한 향상이 포함됩니다. (강 외, 2004), DNA 손상 복구에 관여 (마스토미 외, 2005) 및 RNA 의존성 RNA 중합효소 기능(마이다 외., 2009). 이러한 광범위한 역할에 따라 TERT는 텔로미어뿐만 아니라 염색체를 따라 여러 위치에서 염색질과 연관되어 있는 것으로 밝혀졌습니다. (박 외, 2009,마스토미 외, 2005). 따라서 텔로미어 유지 관리는 TERT가 기여하는 다양한 일련의 기능 중 가장 두드러진 것으로 입증되었습니다. 종양 형성에 대한 텔로머라제의 이러한 추가 기능의 기여는 아직 완전히 밝혀지지 않았습니다.
혈관신생 유도
정상 조직과 마찬가지로 종양에도 영양분과 산소의 형태로 영양분을 공급해야 할 뿐만 아니라 대사 폐기물과 이산화탄소를 배출하는 능력도 필요합니다. 혈관신생 과정에 의해 생성된 종양 관련 신생혈관은 이러한 요구를 해결합니다. 배 발생 동안 혈관계의 발달에는 기존 혈관에서 새로운 혈관이 돋아나는 것(혈관 신생) 외에도 새로운 내피 세포의 탄생과 관으로의 조립(혈관 신생)이 포함됩니다. 이러한 형태형성 이후 정상적인 혈관계는 대체로 정지 상태가 됩니다. 성인에서는 상처 치유 및 여성 생식 순환과 같은 생리적 과정의 일부로 혈관신생이 활성화되지만 일시적으로만 활성화됩니다. 대조적으로, 종양이 진행되는 동안 “혈관신생 스위치”는 거의 항상 활성화되어 유지됩니다. (하나한과 포크맨, 1996).
강력한 증거는 혈관신생 스위치가 혈관신생을 유도하거나 반대하는 상쇄 요인에 의해 지배된다는 것을 나타냅니다. (배리스윌과 크리스토포리, 2009,버거와 벤자민, 2003). 이러한 혈관신생 조절인자 중 일부는 혈관 내피 세포에 의해 표시되는 자극 또는 억제 세포 표면 수용체에 결합하는 신호 전달 단백질입니다. 혈관신생 유도제 및 억제제의 잘 알려진 프로토타입은 각각 혈관 내피 성장 인자-A(VEGF-A) 및 트롬보스폰딘-1(TSP-1)입니다.
VEGF-A 유전자는 배아 및 출생 후 발달 동안 새로운 혈관 성장을 조율하고 내피 세포의 항상성 생존뿐만 아니라 성인의 생리학적 및 병리학적 상황에 관여하는 리간드를 암호화합니다. 3개의 수용체 티로신 키나제(VEGFR-1-3)를 통한 VEGF 신호전달은 이러한 목적의 복잡성을 반영하여 여러 수준에서 조절됩니다. 따라서 VEGF 유전자 발현은 저산소증과 종양유전자 신호전달에 의해 상향조절될 수 있습니다. (페라라, 2009,맥 가반과 포펠, 2008,카르멜리엣, 2005). 추가로, VEGF 리간드는 세포외 기질 분해 프로테아제(예: MMP-9; MMP-9;케센브록 외, 2010). 또한, 섬유아세포 성장 인자(FGF) 계열의 구성원과 같은 다른 혈관신생 신호는 그 발현이 만성적으로 상향조절될 때 종양 혈관신생을 유지하는 것과 관련이 있습니다. (배리스윌과 크리스토포리, 2009). 혈관신생 스위치의 주요 균형점인 TSP-1은 또한 내피 세포에 의해 표시되는 막횡단 수용체와 결합하여 혈관신생 촉진 자극에 대응할 수 있는 억제 신호를 유발합니다.카제로니안 외, 2008).
만성적으로 활성화된 혈관신생과 혈관신생 신호의 불균형한 혼합에 의해 종양 내에서 생성된 혈관은 일반적으로 비정상입니다. 종양 신생혈관은 조숙한 모세혈관 발아, 복잡하고 과도한 혈관 분지, 왜곡되고 확대된 혈관, 불규칙한 혈류, 미세출혈, 누출 및 비정상적인 수준의 내피 세포 증식 및 세포 사멸 (Nagy 외, 2010,발루크 외, 2005).
혈관신생은 동물 모델과 인간 모두에서 침습성 암의 다단계 발달 동안 놀랍게도 초기에 유도됩니다. 다양한 기관에서 발생하는 이형성증 및 상피내 암종을 포함한 전암성 비침습성 병변에 대한 조직학적 분석을 통해 혈관신생 스위치가 조기에 작동하는 것으로 나타났습니다. (Raica 등, 2009,하나한과 포크맨, 1996). 역사적으로 혈관신생은 빠르게 성장하는 육안적 종양이 형성되었을 때만 중요하다고 여겨졌으나, 최근 데이터에 따르면 혈관신생은 종양 진행의 미세한 전암 단계에도 기여하여 암의 필수 특징으로서의 지위를 더욱 확고히 하고 있습니다.
지난 10년 동안 혈관신생에 대한 연구가 놀라울 정도로 쏟아져 나왔습니다. 이러한 풍부한 새로운 지식 속에서 우리는 종양 생리학과 특히 관련된 몇 가지 발전을 강조합니다.
혈관신생 스위치의 변화
일단 혈관신생이 활성화되면, 종양은 혈관신생의 다양한 패턴을 나타냅니다. 췌장 관 선암종과 같은 매우 공격적인 유형을 포함한 일부 종양은 혈관이 저조하고 주로 무혈관이며 실제로 적극적으로 혈관신생을 억제할 수도 있는 간질 “사막”으로 가득 차 있습니다. (올리브 외., 2009). 인간의 신장암과 췌장의 신경내분비암종을 포함한 많은 다른 종양은 혈관신생성이 높고 결과적으로 혈관이 조밀하게 분포되어 있습니다. (지 외., 2010,터너 등, 2003).
종합적으로, 이러한 관찰은 종양 발생 중 혈관신생 스위치의 초기 작동이 진행 중인 혈관신생의 다양한 강도로 이어지며, 후자는 암세포와 관련 간질 미세환경을 모두 포함하는 복잡한 생물학적 가변저항기에 의해 제어된다는 것을 암시합니다. (배리스윌과 크리스토포리, 2009,버거와 벤자민, 2003). 참고로, 최종 결과가 공통 유도 신호(예: VEGF)이더라도 스위칭 메커니즘의 형태는 다양할 수 있습니다. 일부 종양에서는 Ras 및 Myc 와 같은 종양 세포 내에서 작동하는 우세한 종양 유전자가 혈관 신생 인자의 발현을 상향 조절할 수 있는 반면, 다른 종양에서는 이러한 유도 신호가 면역 염증 세포에 의해 간접적으로 생성됩니다. 증식성 신호 전달을 유도하는 종양유전자에 의한 혈관신생의 직접적인 유도는 별개의 특징적 능력이 동일한 형질전환 인자에 의해 공동 조절될 수 있다는 중요한 원리를 보여줍니다.
내인성 혈관신생 억제제는 종양 혈관신생에 자연적인 장벽을 제시합니다
1990년대 연구에서는 TSP-1뿐만 아니라 플라스민(안지오스타틴) 및 18형 콜라겐(엔도스타틴) 조각이 혈관 신생의 내인성 억제제로 작용할 수 있음이 밝혀졌습니다. (리바티, 2009,카제로니안 외, 2008,포크맨, 2006,포크맨, 2002,Nyberg 등, 2005). 지난 10년 동안 그러한 행위자가 12명 더 많이 보고되었습니다(리바티, 2009,포크맨, 2006,Nyberg 등, 2005). 대부분은 단백질이고, 많은 것들은 그 자체로는 혈관신생 조절자가 아닌 구조 단백질의 단백질 분해 절단에 의해 파생됩니다. 이러한 혈관신생의 내인성 억제제 중 상당수는 정상 쥐와 인간의 순환계에서 검출될 수 있습니다. 몇몇 내인성 혈관신생 억제제를 코딩하는 유전자는 불리한 생리학적 효과 없이 마우스 생식계열에서 삭제되었습니다. 그러나 자생 종양과 이식 종양의 성장은 결과적으로 향상됩니다. (리바티, 2009,Nyberg 등, 2005). 대조적으로, 내인성 억제제의 순환 수준이 유전적으로 증가하는 경우(예: 형질전환 마우스 또는 이종 이식 종양의 과발현을 통해) 종양 성장이 손상됩니다. (리바티, 2009,Nyberg 등, 2005); 흥미롭게도, 상처 치유와 지방 축적은 그러한 유전자의 발현이 증가하거나 감소함으로써 손상되거나 가속화됩니다.조(2010),세피넨 외, 2008). 데이터는 이러한 내인성 혈관신생 억제제가 정상적인 상황에서 조직 재형성 및 상처 치유 동안 일시적인 혈관신생을 조절하는 생리적 조절자로서 역할을 한다는 것을 시사합니다. 이는 또한 초기 신생물에 의한 혈관신생의 유도 및/또는 지속에 대한 내인성 장벽으로 작용할 수도 있습니다.
혈관주위세포는 종양 신생혈관의 중요한 구성요소입니다
혈관주위세포는 정상 조직 혈관계에서 내피관의 외부 표면에 밀접하게 인접한 지지 세포로 오랫동안 알려져 왔으며, 여기서 내피 세포에 중요한 기계적 및 생리학적 지지를 제공합니다. 대조적으로, 종양 관련 혈관계는 이들 보조 세포에 의해 감지할 수 있는 범위가 부족한 것으로 묘사되었습니다. 그러나 최근 몇 년간 수행된 주의 깊은 현미경 연구에 따르면 혈관주위세포는 느슨하기는 하지만 모든 종양은 아니더라도 대부분의 신생혈관 구조와 연관되어 있는 것으로 나타났습니다. (라자 외, 2010,버거와 송, 2005). 더 중요한 것은, 아래에 논의된 기계론적 연구를 통해 혈관주위세포 범위가 기능적 종양 신생혈관의 유지에 중요하다는 사실이 밝혀졌습니다.
다양한 골수 유래 세포가 종양 혈관신생에 기여합니다
이제 골수에서 유래하는 세포 유형의 레퍼토리가 병리학적 혈관신생에 중요한 역할을 한다는 것이 분명해졌습니다. (키안과 폴라드, 2010,줌스테그와 크리스토포리, 2009,머독 외, 2008,드 팔마 외, 2007). 여기에는 선천성 면역체계의 세포(특히 대식세포, 호중구, 비만세포, 골수 전구세포)가 포함됩니다. 이들은 전악성 병변과 진행된 종양에 침투하여 그러한 병변의 가장자리에 모입니다. 종양 주위 염증 세포는 이전에 정지 상태였던 조직에서 혈관 신생 스위치를 작동시키고 종양 성장과 관련된 진행 중인 혈관 신생을 유지하는 데 도움을 주며, 아래에 설명된 바와 같이 국소 침습을 촉진합니다. 또한 내피 세포 신호 전달을 표적으로 하는 약물의 효과로부터 혈관계를 보호하는 데 도움이 될 수 있습니다. (페라라, 2010). 또한, 여러 유형의 골수 유래 “혈관 전구 세포”가 특정 사례에서 종양성 병변으로 이동하여 혈관 주위 세포 또는 내피 세포로서 신생 혈관계에 삽입되는 것으로 관찰되었습니다. (Patenaude 외, 2010,코바치치와 뵘, 2009,라마냐와 버거, 2006).
침입 및 전이 활성화
2000년에는 침입과 전이의 기저가 되는 메커니즘이 대체로 수수께끼였습니다. 상피 조직에서 발생하는 암종이 더 높은 병리학적 등급의 악성 종양으로 진행됨에 따라 국소 침윤 및 원격 전이에 반영되며 관련 암세포는 일반적으로 모양뿐만 아니라 다른 세포 및 세포외 기질에 대한 부착에도 변화가 나타납니다. (ECM). 가장 특징적인 변화는 암종 세포에 의한 주요 세포 간 접착 분자인 E-cadherin의 손실과 관련이 있습니다. E-cadherin은 인접한 상피 세포와 부착 접합을 형성함으로써 상피 세포 시트를 조립하고 이러한 시트 내에서 세포의 정지를 유지하는 데 도움을 줍니다. E-cadherin의 발현 증가는 침입과 전이의 길항제로 잘 확립되어 있습니다. 발현의 감소는 이러한 표현형을 강화시키는 것으로 알려져 있습니다. 인간 암종에서 자주 관찰되는 E-cadherin의 하향 조절과 간헐적인 돌연변이 불활성화는 E-cadherin이 이러한 특징적인 능력의 핵심 억제자로서의 역할을 강력하게 뒷받침해 줍니다. (Berx와 반 로이, 2009,카발라로와 크리스토포리, 2004).
추가적으로, 다른 세포-세포 및 세포-ECM 접착 분자를 코딩하는 유전자의 발현은 일부 고도로 공격적인 암종에서 명백히 변경되며, 세포증식을 선호하는 암종은 일반적으로 하향조절됩니다. 반대로, 일반적으로 배아 발생 및 염증 중에 발생하는 세포 이동과 관련된 접착 분자는 종종 상향 조절됩니다. 예를 들어, 기관 형성 과정에서 이동하는 뉴런과 중간엽 세포에서 일반적으로 발현되는 N-cadherin은 많은 침윤성 암종 세포에서 상향 조절됩니다. 이러한 세포-세포/기질 부착 단백질의 획득 및 손실 외에도 침입 및 전이의 주요 조절인자는 대부분 알려지지 않았거나 의심되는 경우 기능적 검증이 부족합니다. (카발라로와 크리스토포리, 2004).
침입과 전이의 다단계 과정은 종종 침입-전이 연속단계(invasion-metasis cascade)라고 불리는 일련의 개별 단계로 도식화되었습니다. (탈매지와 피들러, 2010,피들러, 2003). 이 묘사는 국소적인 침입으로 시작하여 암세포가 인근 혈액 및 림프관으로 침입하고, 림프계 및 조혈계를 통해 암세포가 이동한 다음, 암세포가 이러한 관강의 내강에서 탈출하는 일련의 세포 생물학적 변화를 상상합니다. 혈관이 먼 조직의 실질로 들어가고(유출), 암세포의 작은 결절이 형성되고(미세 전이), 마지막으로 미세 전이 병변이 육안으로 보이는 종양으로 성장합니다. 이 마지막 단계를 “집락화”라고 합니다.
침입과 전이 능력에 대한 연구는 강력하고 새로운 연구 도구와 개선된 실험 모델이 이용 가능해지고 중요한 조절 유전자가 확인됨에 따라 지난 10년 동안 극적으로 가속화되었습니다. 여전히 답을 얻지 못한 주요 질문으로 가득 찬 신흥 분야이지만, 이 복잡한 특징 기능의 중요한 특징을 설명하는 데 상당한 진전이 이루어졌습니다. 이러한 발전에 대한 불완전한 표현은 아래에 강조되어 있습니다.
EMT 프로그램은 침입과 전이를 광범위하게 규제합니다.
“상피-중간엽 전이”(EMT)라고 불리는 발달 조절 프로그램은 변형된 상피 세포가 침입하고, 세포사멸에 저항하고, 전파하는 능력을 획득할 수 있는 수단으로 두드러지게 관련되어 있습니다. (클림코프스키와 사바그너, 2009,폴리악과 와인버그, 2009,티에리 외., 2009,일마즈(Yilmaz)와 크리스토포리(Christofori), 2009년,Barrallo-Gimeno와 Nieto, 2005). 배아의 형태형성과 상처 치유의 다양한 단계에 관여하는 과정을 선택함으로써 암종 세포는 침습과 전이를 가능하게 하는 여러 속성을 동시에 획득할 수 있습니다. 이 다면적인 EMT 프로그램은 침입과 전이 과정에서 암종 세포에 의해 일시적으로 또는 안정적으로, 다양한 정도로 활성화될 수 있습니다.
Snail, Slug, Twist 및 Zeb1/2를 포함한 다발성으로 작용하는 일련의 전사 인자는 배아 발생 동안 EMT 및 관련 이동 과정을 조율합니다. 대부분은 초기에 발달 유전학에 의해 확인되었습니다. 이러한 전사 조절인자는 다수의 악성 종양 유형에서 다양한 조합으로 발현되며 암종 형성의 실험 모델에서 프로그래밍 침입에 인과적으로 중요한 것으로 나타났습니다. 일부는 이소성으로 과발현될 때 전이를 유도하는 것으로 밝혀졌습니다(Micalizzi 외, 2010,타우베 외, 2010,슈말호퍼 외, 2009,양과 와인버그, 2008). 이러한 전사 인자에 의해 유발되는 세포 생물학적 특성에는 부착 접합의 손실 및 다각형 / 상피에서 가느 다란 / 섬유아세포 형태로의 관련 전환, 기질 분해 효소의 발현, 운동성 증가 및 세포사멸에 대한 저항성 증가가 포함됩니다. 모든 특성 침입과 전이 과정에 관여합니다. 이러한 전사 인자 중 몇몇은 E-cadherin 유전자 발현을 직접적으로 억제하여 종양 상피 세포의 운동성과 침습성을 억제하는 주요 인자를 박탈할 수 있습니다. (페이나도 외, 2004).
이용 가능한 증거는 이들 전사 인자가 서로를 조절하고 표적 유전자의 중첩 세트를 조절한다는 것을 시사합니다. 상호 작용과 표현을 지배하는 조건을 설명하는 규칙은 아직 확립되지 않았습니다. 발달 유전학의 증거에 따르면 배아의 이웃 세포로부터 받은 맥락적 신호가 EMT를 통과할 세포에서 이러한 전사 인자의 발현을 촉발하는 데 관여한다는 것이 밝혀졌습니다. (Micalizzi 외, 2010); 유사한 방식으로, 암세포와 인접한 종양 관련 간질 세포의 이형적 상호작용이 이러한 전사 조절인자 중 하나 이상에 의해 조절되는 것으로 알려진 악성 세포 표현형의 발현을 유도할 수 있다는 증거가 늘어나고 있습니다. (카르누브와 와인버그, 2006-2007,브라블츠 외, 2001). 더욱이, 특정 암종의 침윤성 가장자리에 있는 암세포는 EMT를 겪은 것으로 볼 수 있으며, 이는 이러한 암세포가 병변의 중심부에 위치한 암세포가 받는 것과는 다른 미세환경 자극을 받는다는 것을 시사합니다. (Hlubek 등, 2007).
증거는 아직 불완전하지만 EMT 유도 전사 인자는 집락화의 마지막 단계를 제외하고 침입-전이 단계의 대부분의 단계를 조율할 수 있는 것으로 보입니다. 우리는 여전히 EMT에 의해 생성된 중간엽 상태의 다양한 발현과 일시적인 안정성에 대해 거의 알지 못합니다. EMT 유도 전사 인자의 발현이 육종 및 신경외배엽 종양과 같은 특정 비상피 종양 유형에서 관찰되었지만, 이러한 종양에서 악성 특성을 프로그래밍하는 역할은 현재 제대로 문서화되어 있지 않습니다. 또한 침윤성 암종 세포가 EMT 프로그램의 일부 활성화를 통해 반드시 그 능력을 획득하는지, 아니면 대체 규제 프로그램도 이 기능을 활성화할 수 있는지 여부를 결정해야 합니다.
침입 및 전이에 대한 간질 세포의 이형적 기여
암세포와 종양성 간질 세포 사이의 혼선이 침습적 성장과 전이에 대한 획득 능력에 관여한다는 것이 점점 더 분명해지고 있습니다. (Egeblad 등, 2010,키안과 폴라드, 2010,조이스와 폴라드, 2009,칼루리(Kalluri)와 자이스베르그(Zeisberg), 2006년). 이러한 신호는 암종 세포에 영향을 미칠 수 있으며 위에서 제안한 대로 세포의 특징적인 기능을 변경하는 역할을 할 수 있습니다. 예를 들어, 종양 간질에 존재하는 중간엽 줄기세포(MSC)는 암세포가 방출하는 신호에 반응하여 CCL5/RANTES를 분비하는 것으로 밝혀졌습니다. 그런 다음 CCL5는 암세포에 상호 작용하여 침습적 행동을 자극합니다. (Karnoub 등, 2007).
종양 주변의 대식세포는 메탈로프로테이나제 및 시스테인 카텝신 프로테아제와 같은 기질 분해 효소를 공급하여 국소 침입을 촉진할 수 있습니다. (케센브록 외, 2010,조이스와 폴라드, 2009,팔레르모와 조이스, 2008,모하메드와 슬론, 2006); 한 모델 시스템에서는 침입을 촉진하는 대식세포가 암세포에서 생성된 IL-4에 의해 활성화됩니다. (고체바 외, 2010). 그리고 전이성 유방암의 실험 모델에서 종양 관련 대식세포(TAM)는 유방암 세포에 표피 성장 인자(EGF)를 공급하는 반면, 암세포는 CSF-1로 대식세포를 상호 자극합니다. 이들의 공동 상호작용은 순환계로의 혈관내 침입과 암세포의 전이성 확산을 촉진합니다. (키안과 폴라드, 2010,Wyckoff 등, 2007).
이와 같은 관찰은 고등급 악성종양의 표현형이 엄밀하게는 세포 자율적인 방식으로 발생하지 않으며, 그 증상은 종양 세포 게놈 분석을 통해서만 이해할 수 없음을 나타냅니다. 아직 테스트되지 않은 한 가지 중요한 의미는 관련 암세포가 원발성 종양 형성에 필요한 것 이상의 추가 돌연변이를 겪지 않고도 특정 종양에서 침입-전이 연속 단계의 대부분을 협상하는 능력이 획득될 수 있다는 것입니다.
침습적 성장 프로그램의 가소성
침습적 성장 능력(종종 EMT를 통해)을 유도하는 상황별 신호의 역할은 원발 종양에서 더 먼 조직 부위로 전파된 암세포가 더 이상 활성화된 간질 및 침입으로부터 이익을 얻지 못할 수 있다는 점에서 가역성의 가능성을 의미합니다. 원발 종양에 거주하는 동안 경험한 EMT 유도 신호; 이러한 신호에 지속적으로 노출되지 않으면 암종 세포가 새로운 집에서 비침습적 상태로 되돌아갈 수 있습니다. 따라서, 초기 침입 및 전이성 파종 동안 EMT를 겪은 암종 세포는 중간엽-상피 전이(MET)라고 불리는 역과정을 통과할 수 있습니다. (휴고 등, 2007). 더욱이, 암세포가 일상적으로 완전한 EMT 프로그램을 통과한다는 개념은 단순할 가능성이 높습니다. 대신, 많은 경우 암세포는 부분적으로만 EMT 프로그램에 참여할 수 있으므로 잔류 상피 특성을 계속 표현하면서 새로운 중간엽 특성을 획득할 수 있습니다.
다양한 형태의 침입이 다양한 암 유형의 기초가 될 수 있습니다
EMT 프로그램은 “중간엽”이라고 불리는 특정 유형의 침입성을 규제합니다. 또한, 두 가지 다른 뚜렷한 침입 방식이 확인되었으며 암세포 침입과 관련되어 있습니다. (프리들과 울프, 2008,프리들과 울프, 2010). “집단 침입”은 암세포 결절이 인접 조직으로 한꺼번에 전진하는 것과 관련되며, 예를 들어 편평 세포 암종의 특징입니다. 흥미롭게도 이러한 암은 전이되는 경우가 거의 없으므로 이러한 형태의 침윤에는 전이를 촉진하는 특정 기능적 특성이 부족함을 시사합니다. 덜 분명한 것은 “아메바성” 형태의 침입이 널리 퍼져 있다는 것입니다. (매드슨과 사하이, 2010년,사베 외., 2009), 개별 암세포는 형태학적 가소성을 나타내어 중간엽 및 집단 침입 형태 모두에서 발생하는 것처럼 스스로 경로를 확보하기보다는 세포외 기질의 기존 간극을 통해 미끄러져 들어갈 수 있습니다. 집단 및 아메바 형태의 침입에 참여하는 암세포가 EMT 프로그램의 구성 요소를 사용하는지 또는 완전히 다른 세포 생물학적 프로그램이 이러한 대체 침입 프로그램을 구성하는 데 책임이 있는지 여부는 현재 해결되지 않았습니다.
위에서 언급한 또 다른 새로운 개념은 종양 경계에 집합하는 염증 세포에 의한 암세포 침입의 촉진과 관련이 있으며, 세포외 기질 분해 효소와 침습적 성장을 가능하게 하는 기타 요인을 생성합니다. (케센브록 외, 2010,키안과 폴라드, 2010,조이스와 폴라드, 2009); 이러한 기능은 EMT 프로그램의 활성화를 통해 암세포가 이러한 단백질을 생산할 필요성을 제거할 수 있습니다. 따라서 암세포는 기질 분해 효소 자체를 생성하기보다는 전침윤성 염증 세포를 모집하는 화학유인물질을 분비할 수 있습니다.
전이성 집락화의 엄청난 복잡성
전이는 두 가지 주요 단계로 나눌 수 있습니다. 원발 종양에서 먼 조직으로 암세포가 물리적으로 전파되는 단계와 이러한 세포가 외부 조직 미세환경에 적응하여 성공적인 집락화, 즉 미세 전이가 거시적 종양으로 성장하는 단계입니다. 여러 단계의 전파는 EMT 및 유사하게 작용하는 이주 프로그램의 범위에 있는 것으로 보입니다. 그러나 성공적으로 파종되었으나 육안으로 볼 수 있는 전이성 종양으로 결코 진행되지 않는 수많은 미세 전이가 많은 환자에서 입증된 것처럼 집락화는 물리적 파종과 엄격하게 결합되지 않습니다. (탈매지와 피들러, 2010,맥고완 외., 2009,아기레-기소, 2007,타운슨 앤 챔버스, 2006,피들러, 2003).
일부 유형의 암에서는 원발성 종양이 미세 전이를 휴면 상태로 만드는 전신 억제 인자를 분비할 수 있는데, 이는 원발성 성장을 절제한 직후 폭발적인 전이성 성장으로 임상적으로 드러납니다. (데미첼리 외, 2008,포크맨, 2002). 그러나 유방암이나 흑색종과 같은 다른 경우에는 원발 종양이 수술로 제거되거나 약리학적으로 파괴된 후 수십 년 후에 육안으로 전이가 발생할 수 있습니다. 이러한 전이성 종양의 성장은 많은 시행착오 끝에 조직 집락의 복잡한 문제를 해결한 휴면 미세전이를 분명히 반영합니다. (바르칸 외, 2010,아기레-기소, 2007,타운슨 앤 챔버스, 2006).
미세전이에는 혈관신생을 활성화하는 능력과 같이 왕성한 성장에 필요한 다른 특징적인 능력이 부족할 수 있다는 자연사를 추론할 수 있습니다. 실제로 실험적으로 생성된 특정 휴면 미세전이가 육안으로 볼 수 있는 종양을 형성할 수 없는 이유는 종양 혈관신생을 활성화하지 못하기 때문입니다. (나우모프 외, 2008,아기레-기소, 2007). 또한 최근 실험에서는 영양 결핍이 암세포를 수축시키고 가역적 휴면 상태를 채택하게 하는 강력한 자가포식을 유도할 수 있다는 사실이 밝혀졌습니다. 그러한 세포는 더 많은 영양분에 대한 접근과 같은 조직 미세환경의 변화가 허용될 때 이 상태를 종료하고 활발한 성장과 증식을 재개할 수 있습니다. (케니픽 외, 2010,루 외., 2008). 미세전이 휴면의 다른 메커니즘에는 정상 조직 세포외 기질에 내장된 항성장 신호가 포함될 수 있습니다. (바르칸 외, 2010) 및 면역체계의 종양 억제 작용(텡 외, 2008,아기레-기소, 2007).
대부분의 파종된 암세포는 적어도 처음에는 그들이 착륙한 조직의 미세환경에 잘 적응하지 못하는 경향이 있습니다. 따라서 각 유형의 파종된 암세포는 하나 또는 다른 외부 조직의 미세환경에서 번성하는 문제에 대한 자체 임시 솔루션 세트를 개발해야 할 수도 있습니다. (굽타 외, 2005). 이러한 적응에는 수백 가지의 별개의 집락화 프로그램이 필요할 수 있으며, 각 프로그램은 전파되는 암세포의 유형과 집락화가 진행되는 조직 미세환경의 특성에 따라 결정됩니다. 그러나 아래에서 추가로 논의되는 바와 같이, 특정 조직 미세환경은 본질적으로 파종된 암세포에 적합하도록 미리 정해져 있을 수 있습니다. (페이나도 외, 2011,탈매지와 피들러, 2010).
전이성 파종은 오랫동안 다단계 원발성 종양 진행의 마지막 단계로 묘사되어 왔으며 실제로 췌관 선암종의 전이로의 클론 진화에 대한 유전적 증거를 제시하는 최근 게놈 시퀀싱 연구에서 설명된 것처럼 실제로 그럴 가능성이 있는 많은 종양의 경우입니다. (캠벨 외, 2010,뤼벡, 2010,야치다 외, 2010). 반면에, 쥐와 인간 모두에서 세포가 표면적으로 비침습적인 전암성 병변으로부터 분산되어 현저하게 조기에 퍼질 수 있다는 증거가 최근에 나타났습니다. (코글린과 머레이, 2010,클라인, 2009). 추가적으로, 미세전이는 명백히 침습적이지는 않지만 내강 완전성이 결여된 신생혈관을 갖고 있는 원발성 종양으로부터 생성될 수 있습니다. (게르하르트와 셈브, 2008). 암세포는 이러한 종양 전 병변에서 명확하게 전파되고 골수 및 기타 조직에 파종될 수 있지만 이러한 부위에 군집을 형성하고 병리학적으로 중요한 거대 전이로 발전하는 능력은 아직 입증되지 않았습니다. 현재 우리는 이 초기 전이성 전파를 임상적 중요성이 아직 확립되지 않은 생쥐와 인간에서 입증 가능한 현상으로 보고 있습니다.
전파 시기를 넘어서, 암세포가 외부 조직을 거시적 종양으로 정착시키는 능력을 언제, 어디서 발달시키는지도 불분명합니다. 이러한 능력은 전파되기 전 종양의 특정 발달 경로의 결과로 원발성 종양이 형성되는 동안 발생할 수 있으며, 순환계로 유입되는 원발성 종양 세포는 우연히 특정 원격 조직 부위에 정착할 수 있는 능력을 갖게 됩니다. (탈매지와 피들러, 2010). 대안적으로, 특정 조직을 군집화하는 능력은 외부 조직 미세환경에서의 성장에 적응하기 위해 이미 파종된 암세포에 대한 선택적인 압력에 반응하여 발달할 수 있습니다.
이러한 조직 특이적 집락 형성 능력이 발달하면 전이성 집락의 세포는 신체의 새로운 부위뿐만 아니라 조상이 발생한 원발 종양으로 다시 확산될 수 있습니다. 따라서 원발성 종양 내의 세포들 사이에서 명백한 조직 특이적 집락화 프로그램은 원발성 병변 내에서 발생하는 전형적인 종양 진행이 아니라 집으로 돌아온 이민자로부터 유래했을 수 있습니다. (김 외, 2009). 이러한 재파종은 앞서 언급한 인간 췌장암 전이 연구와 일치합니다. (캠벨 외, 2010,뤼벡, 2010,야치다 외, 2010). 다르게 말하면, 원발성 종양 내의 암세포 집단(및 아래에 논의되는 암 줄기세포)의 표현형과 기본 유전자 발현 프로그램은 원격 전이 자손의 역이동에 의해 크게 변형될 수 있습니다.
이 자가 파종 과정에는 또 다른 개념이 내재되어 있습니다. 원발 종양에서 발생하고 악성 특성의 획득에 기여하는 지지 간질은 본질적으로 전이성 병변에서 나오는 순환 암세포에 의한 재파종 및 집락화를 위한 적합한 부위를 제공할 수 있습니다.
전이성 집락화를 가능하게 하는 규제 프로그램을 명확히 하는 것은 향후 연구를 위한 중요한 의제를 나타냅니다. 예를 들어, 특정 조직에서 육안으로 볼 수 있는 전이의 확립과 관련이 있고 이를 촉진하는 것으로 보이는 유전자 세트(“전이성 특징”)를 정의하는 데 상당한 진전이 이루어지고 있습니다. (코글린과 머레이, 2010,보스 등, 2009,올슨 외., 2009,응우옌 외, 2009,굽타 외, 2005). 위에서 언급한 다양한 식민지화 프로그램을 고려할 때 도전 과제는 상당합니다. 더욱이, 식민지화는 세포 자율적 과정에만 전적으로 의존하지 않을 것입니다. 대신, 중요한 간질 지지 세포로 구성된 허용적인 종양 미세 환경의 확립이 거의 확실하게 필요합니다. 이러한 이유로 집락화 과정은 전체적으로 전이성 전파의 이전 단계보다 훨씬 더 복잡하고 다양한 수많은 세포 생물학적 프로그램을 포함할 가능성이 높습니다.
세포내 회로에 의한 인증 기능 프로그래밍
2000년에 우리는 암세포에 영향을 미치는 수많은 신호 분자가 정상 세포에서 작동하는 회로의 재프로그램된 파생물인 정교한 집적 회로의 노드 및 가지로 작동한다는 비유를 제시했습니다. 이후 10년 동안 이러한 회로의 원래 묘사가 확고해졌고 신호 카탈로그와 신호 경로의 상호 연결이 확장되었습니다. 2000년에 이미 그랬던 것처럼 이 회로를 포괄적이고 일관되게 그래픽으로 표현하는 것은 불가능하지는 않더라도 어렵습니다.
이제 우리는 암의 개별 특징과 일치하는 이 회로의 묘사를 제안합니다. 따라서 세포내 집적 회로는 별개의 하위 회로로 분할될 수 있으며, 각 하위 회로는 정상 세포에서 별개의 세포 생물학적 특성을 지원하도록 특화되고 암세포에서 특징적인 기능을 구현하기 위해 재프로그래밍됩니다( 그림 2 ) . 기본 제어 회로에 대한 이해가 부족하거나 여기에 설명된 기능과 광범위하게 겹치기 때문에 이 그림에서는 특징적인 기능의 하위 집합만 다룹니다.
그림 2 세포 내 신호 전달 네트워크는 암세포의 작동을 조절합니다
복잡성의 추가적인 차원에는 상당한 상호 연결과 그에 따른 개별 하위 회로 간의 혼선이 포함됩니다. 예를 들어, 특정 발암성 사건은 돌연변이 RAS 및 상향 조절된 MYC 와 같은 주요 종양 유전자가 여러 특징 기능(예: 증식성 신호 전달, 에너지 대사, 혈관 신생, 침입 및 생존)에 미치는 다양한 효과에서 알 수 있듯이 여러 기능에 영향을 미칠 수 있습니다. 우리는 이 집적 회로의 향후 버전이 아직 여기서 다루지 않은 하위 회로 및 관련 특징 기능을 포함할 것으로 예상합니다.
특성 및 새로운 특징 활성화
우리는 암세포가 생존하고, 증식하고, 퍼질 수 있도록 하는 획득된 기능적 능력으로 암의 특징을 정의했습니다. 이러한 기능은 다양한 종양 유형에서 뚜렷한 메커니즘을 통해 그리고 다단계 종양 형성 과정 중 다양한 시점에 획득됩니다. 이들의 획득은 두 가지 가능 특성에 의해 가능해집니다. 가장 두드러진 것은 염색체 재배열을 포함한 무작위 돌연변이를 생성하는 암세포의 게놈 불안정성의 발달입니다. 그중에는 특징적인 능력을 조율할 수 있는 희귀한 유전적 변화가 있습니다. 두 번째 가능하게 하는 특징은 면역 체계의 세포에 의해 유발되는 전악성 및 솔직히 악성 병변의 염증 상태와 관련되며, 그 중 일부는 다양한 수단을 통해 종양 진행을 촉진하는 역할을 합니다.
그러나 암세포의 다른 뚜렷한 특성은 암 발병에 기능적으로 중요한 것으로 제안되었으며 따라서 핵심 특징 목록에 추가될 수 있습니다. (네그리니 외, 2010,루오 외., 2009,콜로타 외., 2009). 이러한 두 가지 속성은 특히 매력적입니다. 첫 번째는 지속적인 세포 성장과 증식을 지원하기 위해 세포 에너지 대사의 주요 재프로그래밍을 포함하며, 대부분의 정상 조직에서 작동하고 관련 세포의 생리학적 작동에 연료를 공급하는 대사 프로그램을 대체합니다. 두 번째는 암세포가 공격을 적극적으로 회피하고 면역세포에 의한 제거를 포함합니다. 이 기능은 종양 발달과 진행을 길항하고 강화하는 면역 체계의 이분법적인 역할을 강조합니다. 이러한 능력은 모두 다양한 형태의 인간 암의 발달과 진행을 촉진하는 것으로 입증될 수 있으므로 암의 새로운 특징으로 간주될 수 있습니다. 이러한 활성화 특성과 새로운 특징은 다음과 같습니다 .그림 3 은 아래에서 개별적으로 설명됩니다.
그림 3 새로운 특징 및 활성화 특성
활성화 특성: 게놈 불안정성과 돌연변이
위에 열거된 여러 특징의 획득은 종양 세포 게놈의 연속적인 변경에 크게 좌우됩니다. 간단히 설명하면 특정 돌연변이 유전자형은 세포의 하위 클론에 선택적인 이점을 부여하여 국소 조직 환경에서 파생물과 최종 우세를 가능하게 합니다. 따라서, 다단계 종양 진행은 연속적인 클론 확장으로 묘사될 수 있으며, 각각은 가능하게 하는 돌연변이 유전자형의 우연한 획득에 의해 촉발됩니다. 종양 억제 유전자의 불활성화와 같은 유전적 표현형은 DNA 메틸화 및 히스톤 변형과 같은 후생적 메커니즘을 통해 획득될 수도 있기 때문입니다. (베르다스코와 에스텔러, 2010,에스텔러, 2007,존스와 베일린, 2007), 일부 클론 확장은 유전자 발현 조절에 영향을 미치는 비돌연변이 변화에 의해 촉발될 수 있습니다.
DNA의 결함을 감지하고 해결하는 게놈 유지 시스템의 뛰어난 능력은 각 세포 생성 중에 자연 돌연변이 비율이 일반적으로 매우 낮다는 것을 보장합니다. 종양 형성을 조율하는 데 필요한 돌연변이 유전자 목록을 획득하는 과정에서 암세포는 종종 돌연변이 비율을 증가시킵니다.네그리니 외, 2010,솔크 외, 2010). 이러한 돌연변이성은 돌연변이 유발 물질에 대한 민감도 증가, 게놈 유지 관리 기계의 하나 또는 여러 구성 요소의 고장, 또는 두 가지 모두를 통해 달성됩니다. 또한 일반적으로 게놈 무결성을 모니터링하고 유전적으로 손상된 세포를 노화 또는 세포사멸로 만드는 감시 시스템을 손상시킴으로써 돌연변이 축적이 가속화될 수 있습니다. (잭슨과 바텍, 2009,카스탄, 2008,시갈과 로터, 2000). TP53의 역할은 여기서 핵심적이며, 이를 “게놈의 수호자”라고 부르게 됩니다(레인, 1992).
종종 게놈의 “관리자”라고 불리는 DNA 유지 관리 기계의 다양한 구성 요소에 영향을 미치는 다양한 결함(킨즐러(Kinzler)와 보겔슈타인(Vogelstein), 1997년)—문서화되었습니다. 이러한 관리인 유전자의 결함 목록에는 (1) DNA 손상을 감지하고 복구 기계를 활성화하고, (2) 손상된 DNA를 직접 복구하고, (3) DNA가 손상되기 전에 돌연변이 유발 분자를 비활성화하거나 차단하는 제품이 포함됩니다. (네그리니 외, 2010,치시아와 엘리지, 2010,잭슨과 바텍, 2009,카스탄, 2008,하퍼와 엘리지, 2007,프리드버그 외, 2006). 유전적 관점에서 볼 때 이러한 관리인 유전자는 종양 억제 유전자와 매우 유사하게 행동합니다. 즉, 종양이 진행되는 동안 그 기능이 손실될 수 있으며, 이러한 손실은 돌연변이 비활성화 또는 후생적 억제를 통해 달성됩니다. 이들 관리인 유전자 중 상당수의 돌연변이 복사본이 쥐의 생식계열에 도입되어 예상대로 암 발생률이 증가하고 인간의 암 발병에 잠재적으로 관여할 가능성이 있음을 뒷받침합니다. (반스와 린달, 2004년).
우리가 처음 암의 특징을 열거한 이후 10년 동안 종양과 관련된 게놈 불안정성의 또 다른 주요 원인이 밝혀졌습니다. 앞서 설명한 것처럼 많은 종양에서 텔로미어 DNA의 손실은 핵형 불안정성과 관련 증폭 및 염색체 부분의 삭제를 유발합니다. (아르탄디와 드피뉴, 2010). 이러한 관점에서 볼 때, 텔로머라제는 무제한 복제 잠재력을 위한 특징적인 기능을 가능하게 하는 것 이상이며 게놈 무결성을 유지하는 중요한 관리인 목록에 추가되어야 합니다.
암세포 게놈에 대한 분자유전학적 분석의 발전은 기능을 바꾸는 돌연변이와 종양 진행 중 진행 중인 게놈 불안정성에 대한 가장 설득력 있는 증거를 제공했습니다. 한 가지 분석 유형인 비교 게놈 혼성화(CGH)는 세포 게놈 전반에 걸쳐 유전자 카피 수의 증가와 손실을 문서화합니다. 많은 종양에서 CGH에 의해 밝혀진 광범위한 게놈 이상은 게놈 무결성 제어 상실에 대한 명확한 증거를 제공합니다. 중요한 것은 게놈의 특정 부위에서 특정 이상(증폭 및 결실 모두)이 재발한다는 것은 그러한 부위에 신생물성 진행을 선호하는 변형이 있는 유전자가 있을 가능성이 있음을 나타냅니다. (코르콜라와 그레이, 2010).
최근에는 효율적이고 경제적인 DNA 서열분석 기술의 출현으로 고해상도 분석이 가능해졌습니다. 초기 연구에서는 다양한 종양 유형에서 독특한 DNA 돌연변이 패턴이 밝혀지고 있습니다( http://cancergenome.nih.gov/ 참조 ). 그리 멀지 않은 미래에 전체 암세포 게놈의 서열 분석은 암세포 게놈 전체에 흩어져 있는 표면상 무작위 돌연변이의 유병률을 명확히 할 것을 약속합니다. 따라서 반복되는 유전적 변화는 종양 발병기전에서 특정 돌연변이의 인과적 역할을 가리킬 수 있습니다.
게놈 변경의 세부 사항은 종양 유형에 따라 극적으로 다르지만 인간 종양에서 이미 기록된 수많은 게놈 유지 및 복구 결함과 유전자 카피 수 및 뉴클레오티드 서열의 광범위한 불안정화에 대한 풍부한 증거는 우리에게 불안정성을 확신시킵니다. 게놈의 대부분은 인간 암세포에 내재되어 있습니다. 이는 결과적으로 게놈 유지 및 복구의 결함이 선택적으로 유리하며 따라서 종양 진행에 도움이 된다는 결론으로 이어집니다. 이는 진화하는 전암 세포가 유리한 유전자형을 축적할 수 있는 속도를 가속화하기 때문입니다. 따라서 게놈 불안정성은 특징적인 능력 획득과 인과적으로 연관되는 활성화 특성임이 분명합니다.
활성화 특성: 종양 촉진 염증
병리학자들은 일부 종양이 면역계의 선천적 및 적응적 부분의 세포에 의해 조밀하게 침윤되어 비종양성 조직에서 발생하는 염증 상태를 반영한다는 것을 오랫동안 인식해 왔습니다. (드보르작, 1986). 면역체계의 개별 세포 유형을 정확하게 식별하기 위한 더 나은 마커의 출현으로 이제 사실상 모든 종양 병변에는 세포 유형별 항체로만 감지할 수 있는 미묘한 침윤부터 심각한 염증에 이르기까지 다양한 밀도로 존재하는 면역 세포가 포함되어 있다는 것이 분명해졌습니다. 표준 조직화학적 염색 기술로도 명백하게 나타납니다(페이지 외., 2010). 역사적으로 이러한 면역 반응은 주로 종양을 근절하려는 면역계의 시도를 반영하는 것으로 생각되었으며, 실제로 아래에 논의된 바와 같이 면역 파괴를 회피하도록 종양에 압력을 가하면서 많은 종양 유형에 대한 항종양 반응에 대한 증거가 증가하고 있습니다.
2000년에는 종양 관련 염증 반응이 종양 형성 및 진행을 향상시키는 예상치 못한 역설적인 효과를 가져 사실상 초기 종양이 특징적인 능력을 획득하는 데 도움이 된다는 단서가 이미 나타났습니다. 이후 10년 동안 염증과 암 병인 사이의 교차점에 대한 연구가 꽃피웠고, 주로 선천성 면역 체계의 면역 세포가 종양 진행에 미치는 기능적으로 중요한 종양 촉진 효과에 대한 풍부하고 설득력 있는 입증이 이루어졌습니다. (DeNardo 외, 2010,그리벤니코프 외, 2010,키안과 폴라드, 2010,콜로타 외., 2009). 염증은 증식성 신호 전달을 유지하는 성장 인자, 세포 사멸을 제한하는 생존 인자, 혈관신생 인자, 혈관 신생, 침입 및 전이를 촉진하는 세포외 기질 변형 효소, 유도성 등의 생체 활성 분자를 종양 미세환경에 공급함으로써 다양한 특징에 기여할 수 있습니다. EMT 및 기타 특징 촉진 프로그램의 활성화로 이어지는 신호(DeNardo 외, 2010,그리벤니코프 외, 2010,키안과 폴라드, 2010,카르누브와 와인버그, 2006-2007).
중요한 것은 염증이 어떤 경우에는 종양 진행의 초기 단계에서 분명하게 나타나고 초기 종양이 본격적인 암으로 발전하는 것을 촉진할 수 있다는 것입니다. (키안과 폴라드, 2010,드 Visser 외, 2006). 또한, 염증 세포는 인근 암세포에 적극적으로 돌연변이를 일으키는 화학 물질, 특히 활성 산소종을 방출하여 악성 종양 상태로의 유전적 진화를 가속화할 수 있습니다. (그리벤니코프 외, 2010). 따라서 염증은 핵심 특징 기능 획득에 기여하는 활성화 특성으로 간주될 수 있습니다. 이러한 활성화 특성을 담당하는 세포는 종양 미세 환경에 대한 아래 섹션에 설명되어 있습니다.
새로운 특징: 에너지 대사 재프로그래밍
신생물성 질환의 본질을 나타내는 만성적이고 흔히 통제되지 않는 세포 증식은 세포 증식의 규제 완화된 조절뿐만 아니라 세포 성장과 분열을 촉진하기 위한 에너지 대사의 상응하는 조정도 포함합니다. 호기성 조건에서 정상 세포는 포도당을 처리하여 먼저 세포질에서 해당과정을 통해 피루브산으로, 그 다음 미토콘드리아에서 이산화탄소로 전환합니다. 혐기성 조건에서는 해당과정이 선호되고 상대적으로 적은 양의 피루브산이 산소를 소비하는 미토콘드리아로 보내집니다. Otto Warburg는 처음으로 암세포 에너지 대사의 변칙적인 특성을 관찰했습니다. (바르부르크, 1930년,워버그, 1956a,워버그, 1956b): 산소가 있는 경우에도 암세포는 에너지 대사를 주로 해당작용으로 제한하여 “호기성 해당작용”이라고 불리는 상태로 이어짐으로써 포도당 대사와 그에 따른 에너지 생산을 재프로그램할 수 있습니다.
암세포에서 이러한 대사 스위치의 존재는 이후 수십 년 동안 입증되었습니다. 에너지 대사의 이러한 재프로그램화는 암세포가 미토콘드리아 산화적 인산화에 비해 해당과정에 의해 제공되는 ATP 생산의 약 18배 낮은 효율을 보상해야 한다는 점에서 겉으로는 직관에 어긋나는 것 같습니다. 그들은 부분적으로 포도당 운반체, 특히 GLUT1을 상향 조절하여 세포질로의 포도당 유입을 실질적으로 증가시킵니다. (존스와 톰슨, 2009,DeBerardinis 외, 2008,수와 사바티니, 2008). 실제로, 포도당의 현저하게 증가된 흡수 및 활용은 많은 인간 종양 유형에서 기록되었으며, 가장 쉽게 보고자로서 방사선 표지된 포도당 유사체( 18F- 플루오로데옥시글루코스, FDG)를 사용하여 양전자 방출 단층 촬영(PET)을 사용하여 포도당 흡수를 비침습적으로 시각화함으로써 가장 쉽게 기록되었습니다.
해당과정의 연료 공급은 활성화된 종양유전자(예: RAS , MYC ) 및 돌연변이 종양 억제인자(예: TP53 ) 와 연관되어 있는 것으로 나타났습니다. (DeBerardinis 외, 2008,존스와 톰슨, 2009), 종양 세포의 변경은 주로 세포 증식의 특징적인 능력을 부여하고 세포 증식 억제 조절을 피하며 세포사멸을 약화시키는 이점 때문에 선택되었습니다. 해당과정에 대한 이러한 의존성은 많은 종양 내에서 작동하는 저산소 조건 하에서 더욱 강조될 수 있습니다. 저산소증 반응 시스템은 포도당 운반체와 해당 과정의 여러 효소를 상향 조절하기 위해 다발성 작용을 합니다. (세멘자, 2010a,존스와 톰슨, 2009,DeBerardinis 외, 2008). 따라서 Ras 종양단백질과 저산소증은 모두 HIF1α와 HIF2α 전사 인자의 수준을 독립적으로 증가시킬 수 있으며, 이는 차례로 해당과정을 상향 조절합니다. (세멘자, 2010a,세멘자, 2010b,Kroemer와 Pouyssegur, 2008).
암세포의 해당과정 스위치에 대한 기능적 근거는 미토콘드리아 산화적 인산화에 비해 해당과정에 의한 ATP 생성 효율이 상대적으로 낮기 때문에 파악하기 어렵습니다. 오랫동안 잊혀진 한 사람에 따르면 (포터, 1958) 그리고 최근 부활하고 정제된 가설(밴더 하이덴 외, 2009), 증가된 해당작용은 해당과정 중간체를 뉴클레오시드 및 아미노산 생성 경로를 포함한 다양한 생합성 경로로 전환시킬 수 있습니다. 이는 새로운 세포를 조립하는 데 필요한 거대분자와 세포소기관의 생합성을 촉진합니다. 더욱이, Warburg와 같은 대사는 빠르게 분열하는 많은 배아 조직에 존재하는 것으로 보이며, 이는 활성 세포 증식에 필요한 대규모 생합성 프로그램을 지원하는 역할을 다시 한번 시사합니다.
흥미롭게도 일부 종양에는 에너지 생성 경로가 다른 두 개의 암세포 하위 집단이 포함되어 있는 것으로 밝혀졌습니다. 한 하위 집단은 젖산을 분비하는 포도당 의존성(“바르부르크 효과”) 세포로 구성되는 반면, 두 번째 하위 집단의 세포는 구연산 주기의 일부를 사용하여 이웃이 생산한 젖산염을 주요 에너지원으로 우선적으로 수입하고 활용합니다. 그래서 (케네디와 드워스트, 2010,페론, 2009,세멘자, 2008). 이 두 개체군은 분명히 공생적으로 기능합니다. 저산소 암세포는 포도당을 연료로 사용하고 젖산을 폐기물로 분비합니다. 젖산은 더 잘 산소화된 형제에 의해 수입되어 우선적으로 연료로 사용됩니다. 이러한 종양 내 공생의 도발적인 방식은 아직 일반화되지 않았지만, 종양 성장을 촉진하기 위한 젖산 분비 세포와 젖산 활용 세포 간의 협력은 실제로 종양의 발명이 아니라 오히려 다시 정상적인 생리적 메커니즘의 결합을 반영합니다. 근육에서 작동하는 것(케네디와 드워스트, 2010,페론, 2009,세멘자, 2008). 또한, 정상산소증에서 저산소증에 이르는 산소 공급은 종양에서 반드시 정적인 것이 아니라 일시적으로나 지역적으로 변동한다는 것이 분명해지고 있습니다. (하디 외, 2009), 종양 관련 신생혈관의 불안정성과 혼란스러운 조직의 결과일 가능성이 높습니다.
변화된 에너지 대사는 암의 특징으로 받아들여지는 다른 많은 암 관련 특성만큼 암세포에도 널리 퍼져 있는 것으로 입증되었습니다. 이러한 깨달음은 세포 에너지 대사 조절을 완화하는 것이 잘 확립된 6가지 핵심 특징만큼 근본적인 암세포의 핵심 특징 기능인지 여부에 대한 의문을 제기합니다. 실제로, 에너지 대사의 방향 전환은 주로 암의 핵심 특징을 프로그래밍하는 데 어떤 방식으로든 관여하는 단백질에 의해 조정됩니다. 이러한 방식으로 볼 때, 호기성 해당작용은 증식을 유도하는 종양유전자에 의해 프로그램되는 또 다른 표현형일 뿐입니다.
흥미롭게도 IDH(isocitrate dehydrogenase 1/2) 효소의 활성화(기능 획득) 돌연변이가 신경아교종 및 기타 인간 종양에서 보고되었습니다. (엔 등, 2010). 이러한 돌연변이가 에너지 대사를 변경하는 능력으로 인해 클론적으로 선택된 것으로 입증될 수 있지만, 이들의 활성이 HIF-1 전사 인자의 산화 및 안정성 증가와 연관되어 있다는 혼란스러운 데이터가 있습니다. (라이트먼과 얀, 2010) 이는 각각 게놈 안정성과 혈관 신생/침습에 영향을 미쳐 표현형 경계선이 모호해질 수 있습니다. 따라서 현재 재프로그램된 에너지 대사를 신흥 특징으로 지정하는 것은 그것의 명백한 중요성뿐만 아니라 핵심 특징으로부터의 기능적 독립성을 둘러싼 해결되지 않은 문제를 강조하기 위해 가장 적절해 보입니다.
새로운 특징: 면역 파괴 회피
종양 형성을 둘러싼 아직 해결되지 않은 두 번째 문제는 초기 종양, 말기 종양 및 미세 전이의 형성과 진행을 저항하거나 근절하는 데 면역계가 수행하는 역할과 관련이 있습니다. 면역 감시에 대한 오랜 이론은 세포와 조직이 항상 경계하는 면역 체계에 의해 지속적으로 모니터링되고, 그러한 면역 감시가 대다수의 초기 암세포와 초기 종양을 인식하고 제거하는 역할을 한다고 제안합니다. 이 논리에 따르면, 나타나는 고형 종양은 어떻게든 면역 체계의 다양한 부문에 의한 탐지를 피하거나 면역학적 살해 범위를 제한하여 박멸을 피할 수 있었습니다.
종양에 대한 결함 있는 면역학적 모니터링의 역할은 면역이 저하된 개인에게서 특정 암이 눈에 띄게 증가하는 것으로 입증되는 것 같습니다. (바이디치(Vajdic)와 반 레이우웬(van Leeuwen), 2009년). 그러나 이들 중 대다수는 바이러스에 의해 유발된 암이며, 이는 이러한 종류의 암에 대한 대부분의 통제가 일반적으로 부분적으로 바이러스에 감염된 세포를 제거함으로써 감염된 개인의 바이러스 부담을 줄이는 데 달려 있음을 시사합니다. 따라서 이러한 관찰은 비바이러스성 병인의 종양의 80% 이상 형성을 제한하는 데 있어 면역계의 가능한 역할에 대해 거의 밝히지 못하는 것으로 보입니다. 그러나 최근 몇 년 동안 유전적으로 조작된 쥐와 임상 역학 모두에서 점점 더 많은 증거가 나오고 있는데, 이는 적어도 비바이러스로 유발된 암의 일부 형태에서 면역 체계가 종양 형성과 진행에 대한 중요한 장벽으로 작용한다는 사실을 시사합니다.
면역 체계의 다양한 구성 요소가 결핍되도록 유전적으로 조작된 쥐를 발암 물질 유발 종양의 발달에 대해 평가했을 때, 면역 능력이 있는 대조군에 비해 면역 결핍 쥐에서 종양이 더 자주 발생하고/하거나 더 빠르게 자라는 것이 관찰되었습니다. 특히, CD8 + 세포독성 T 림프구(CTL), CD4 + T h 의 발달 또는 기능 결함1개의 보조 T 세포 또는 자연 살해(NK) 세포는 각각 종양 발생률의 입증 가능한 증가를 가져왔습니다. 더욱이, T 세포와 NK 세포 모두에 복합적인 면역결핍이 있는 쥐는 암 발병에 훨씬 더 취약했습니다. 결과는 적어도 특정 실험 모델에서 면역 체계의 선천적 및 적응적 세포 부문이 모두 면역 감시 및 그에 따른 종양 근절에 크게 기여할 수 있음을 나타냅니다. (텡 외, 2008,김 외, 2007).
또한, 이식 실험에 따르면 원래 면역 결핍 쥐에서 발생한 암세포는 동계 면역 능력이 있는 숙주에서 이차 종양을 시작하는 데 종종 비효율적인 반면, 면역 능력이 있는 쥐에서 발생한 종양의 암세포는 두 유형의 숙주 모두에서 이식된 종양을 시작하는 데 동등하게 효율적입니다.텡 외, 2008,김 외, 2007). 이러한 행동은 다음과 같이 해석되었습니다. 면역 능력이 있는 숙주에서는 면역원성이 높은 암세포 클론이 일상적으로 제거됩니다. 이 과정을 “면역 편집”이라고 합니다. 면역원성이 약한 변이체만 남기고 성장하여 고형 종양이 생성됩니다. 그런 약한 면역원성 세포는 이후에 면역결핍 숙주와 면역능력이 있는 숙주 모두에 군집을 형성할 수 있습니다. 반대로, 면역결핍 숙주에서 발생하는 경우, 면역원성 암세포는 선택적으로 고갈되지 않고 대신 면역원성이 약한 세포와 함께 번성할 수 있습니다. 이러한 편집되지 않은 종양의 세포가 동계 수용자에게 연속적으로 이식되면 면역원성 암세포는 처음으로 2차 숙주의 유능한 면역 체계에 직면했을 때 거부됩니다. (스미스 외., 2006). (이러한 특정 실험에서는 그러한 종양을 유발하는 데 사용된 화학적 발암 물질이 특히 면역원성을 갖는 암세포를 생성하는 경향이 있는지 여부에 대한 질문에 답이 없습니다.)
임상 역학은 또한 인간 암의 일부 형태에서 항종양 면역 반응의 존재를 점점 더 뒷받침하고 있습니다. (Bindea 등, 2010,페론과 드라노프, 2010,넬슨, 2008). 예를 들어, CTL과 NK 세포가 심하게 침윤된 결장 및 난소 종양 환자는 풍부한 킬러 림프구가 부족한 환자보다 예후가 더 좋습니다. (페이지 외., 2010,넬슨, 2008); 다른 암에 대한 사례는 시사적이지만 덜 설득력이 있으며 지속적인 조사 대상입니다. 또한, 면역이 억제된 일부 장기 이식 수혜자에게서 기증자 유래 암이 발생하는 것이 관찰되었는데, 이는 표면상 종양이 없는 기증자에게서 암세포가 완전히 기능하는 면역 체계에 의해 휴면 상태에서 억제되고 있음을 시사합니다. (스트라우스와 토마스, 2010).
그럼에도 불구하고, 만성 면역억제 환자의 역학은 위에서 언급한 바와 같이 주요 형태의 비바이러스성 인간 암의 발생률이 유의하게 증가했음을 나타내지 않습니다. 이는 종양 형성 및 종양 진행에 대한 효과적인 장벽으로서 면역 감시의 중요성에 대한 논쟁으로 간주될 수 있습니다. 그러나 우리는 HIV와 약리학적으로 면역이 억제된 환자는 주로 T 및 B 세포 구획에서 면역결핍을 나타내므로 NK 세포와 CTL이 모두 결여된 유전적으로 조작된 돌연변이 쥐에서 생성된 다성분 면역학적 결핍을 나타내지 않습니다. 이는 그러한 환자가 NK 및 기타 선천성 면역 세포에 의해 탑재된 암에 대한 면역학적 방어에 대한 잔여 능력을 여전히 가지고 있을 가능성을 열어줍니다.
사실, 암 면역학에 대한 위의 논의는 종양-숙주 면역학적 상호작용을 단순화합니다. 왜냐하면 면역원성이 높은 암세포는 이를 제거하기 위해 파견된 면역 체계의 구성 요소를 비활성화함으로써 면역 파괴를 피할 수 있기 때문입니다. 예를 들어, 암세포는 TGF-β 또는 기타 면역억제 인자를 분비하여 침윤된 CTL 및 NK 세포를 마비시킬 수 있습니다. (양 외, 2010,쉴즈 외, 2010). 보다 미묘한 메커니즘은 조절 T 세포(Treg) 및 골수 유래 억제 세포(MDSC)를 포함하여 적극적으로 면역을 억제하는 염증 세포의 모집을 통해 작동합니다. 둘 다 세포독성 림프구의 활동을 억제할 수 있습니다. (무지아카코스 외, 2010,오스트랜드-로젠버그와 신하, 2009).
이러한 고려 사항과 인간의 종양 형성 및 진행에 대한 중요한 장벽인 항종양 면역의 아직 초보적인 입증을 고려하여 우리는 면역 회피를 또 다른 새로운 특징으로 제시하며, 핵심 특징 기능으로서의 일반성은 확고히 확립되어야 합니다.
종양 미세환경
지난 10년 동안 종양은 정상적인 건강한 조직의 복잡성에 근접하거나 심지어 그 수준을 초과할 수도 있는 기관으로 점차 인식되어 왔습니다. 이러한 관점에서 볼 때, 종양의 생물학은 그 안에 있는 개별 특수 세포 유형( 그림 4 , 위쪽)과 다단계 종양 형성 과정에서 구성되는 “종양 미세 환경”을 연구함으로써만 이해할 수 있습니다 ( 그림 4, 낮추다). 이러한 묘사는 종양을 상대적으로 균질한 암세포의 집합에 불과하며, 이러한 세포의 세포 자율적 특성을 밝혀냄으로써 암세포의 전체 생물학을 이해할 수 있다는 이전의 환원주의적 관점과 완전히 대조됩니다. 우리는 여기에서 많은 종양의 생물학에 중요한 방식으로 기여하는 것으로 알려진 일련의 세포 유형을 열거하고 개별 및 집단 기능을 제어하는 조절 신호에 대해 논의합니다. 이러한 관찰의 대부분은 신생물 상피 세포가 종양 관련 간질을 형성하는 중간엽 세포와 분명히 구별되는 구획(실질)을 구성하는 암종 연구에서 비롯됩니다.
그림 4 종양 미세환경의 세포
암세포와 암줄기세포
암세포는 질병의 기초입니다. 그들은 종양을 시작하고 종양 진행을 촉진하여 암을 유전 질환으로 정의하는 발암성 및 종양 억제 돌연변이를 운반합니다. 전통적으로, 종양 내의 암세포는 종양 진행 과정의 상대적으로 늦은 시점까지 합리적으로 균질한 세포 집단으로 묘사되어 왔으며, 증가된 유전적 불안정성과 결합된 과다증식으로 인해 별개의 클론 하위 집단이 생성됩니다. 이러한 클론 이질성을 반영하여 많은 인간 종양은 다양한 정도의 분화, 증식, 혈관성, 염증 및/또는 침습성에 의해 구분되는 영역을 포함하여 조직병리학적으로 다양합니다. 그러나 최근에는 증거는 여전히 단편적이지만, CSC는 비록 매우 다양하게 존재하지만 대부분은 아니더라도 많은 종양의 공통 구성 요소임이 입증될 수 있습니다. CSC는 수용 숙주 마우스에 접종 시 새로운 종양을 효율적으로 파종하는 능력을 통해 운영상 정의됩니다. (조와 클라크, 2008,로보 등, 2007). 이러한 기능적 정의는 종종 기원 조직의 정상 줄기 세포에서도 발현되는 마커의 CSC에서의 발현을 포함함으로써 보완됩니다. (알-하지 외, 2003).
CSC는 초기에 조혈 악성종양의 발병에 연루되어 있었습니다. (레야 외., 2001,보닛과 딕, 1997) 그리고 몇 년 후 고형 종양, 특히 유방암종과 신경외배엽 종양에서 확인되었습니다(길버트슨과 리치, 2007,알-하지 외, 2003). 표시된 세포 표면 마커를 기반으로 암세포를 분별하면 면역결핍 마우스에 이식 시 새로운 종양을 심는 능력이 해당 대다수 집단에 비해 크게 향상된 신생물 세포 하위 집단이 생성되었습니다. 이러한 희귀한 종양 개시 세포는 특정 정상 조직 줄기 세포 집단과 전사 프로필을 공유하는 것으로 입증되어 줄기 유사 세포로 지정되었습니다.
고형 종양 내 CSC의 기원은 명확하지 않으며 실제로 종양 유형마다 다를 수 있습니다. 일부 종양에서는 정상 조직 줄기세포가 발암성 변형을 거쳐 CSC를 생성하는 기원 세포 역할을 할 수 있습니다. 다른 경우에는 부분적으로 분화된 통과 증폭 세포(전조 세포라고도 함)가 초기 발암성 형질전환을 겪은 후 줄기와 유사한 특성을 갖게 될 수 있습니다. 일단 원발성 종양이 형성되면, CSC는 정상적인 종양과 마찬가지로 자가 재생될 뿐만 아니라 더욱 분화된 파생물을 생성할 수 있습니다. 종양성 CSC의 경우 이러한 후손 세포는 많은 종양의 대부분을 형성합니다. 종양이 시작되고 이후의 다단계 진행 중에 점점 증가하는 신생물성 줄기 세포의 여러 가지 별개의 클래스가 형성되는지 여부는 아직 확립되지 않았습니다.
최근 연구에서는 CSC 특성 획득과 위에서 논의한 EMT 전환분화 프로그램을 상호 연관시켰습니다. (싱과 정착민, 2010,마니 등, 2008,모렐 외., 2008). 특정 모델 시스템에서 이 프로그램을 유도하면 자가 재생 능력 및 정상 줄기 세포와 암 줄기 세포 모두와 관련된 항원 표현형을 포함하여 줄기 세포의 많은 정의 기능을 유도할 수 있습니다. 이러한 일치는 EMT 프로그램이 암세포가 원발성 종양에서 물리적으로 전파될 수 있게 할 뿐만 아니라 전파 부위에서 후속 클론 확장에 결정적인 자가 재생 능력을 그러한 세포에 부여할 수 있음을 시사합니다. (브라블츠 외, 2005). 일반화하면 이 연결은 중요한 추론 가설을 제기합니다. 활성화된 염증성 간질에 의해 방출되는 신호와 같은 EMT를 유발하는 이형 신호는 CSC를 생성하고 유지하는 데에도 중요할 수 있습니다.
점점 더 많은 수의 인간 종양이 마우스에 이종 이식 시 효율적인 종양 개시 능력을 통해 작동적으로 정의된 CSC의 특성을 가진 하위 집단을 포함하는 것으로 보고되었습니다. 그럼에도 불구하고 종양 세포의 뚜렷한 표현형 하위 클래스로서 CSC의 중요성은 종양 내에서 자주 인용되는 희귀성과 마찬가지로 논쟁의 문제로 남아 있습니다. (보이코 외, 2010,굽타 외, 2009,퀸타나 외., 2008). 실제로, 종양 내에서 작용하는 표현형 가소성은 CSC와 비-CSC 사이의 양방향 상호전환을 생성하여 CSC의 상대적인 풍부함에 동적인 변화를 가져올 수 있다는 것이 타당합니다. 이러한 가소성은 유병률에 대한 확실한 측정을 복잡하게 만들 수 있습니다. 유사한 가소성은 이미 가역적으로 참여할 수 있는 EMT 프로그램에 관련되어 있습니다. (티에리와 슬리먼, 2006).
이러한 복잡성에도 불구하고, 종양 이질성의 새로운 차원이 성공적인 암 치료법에 중요한 영향을 미친다는 것은 분명합니다. 다양한 종양 유형에서 증가하는 증거는 CSC의 특성을 가진 세포가 일반적으로 사용되는 다양한 화학요법 치료에 더 저항성이 있음을 시사합니다. (싱과 정착민, 2010,크레이튼 외, 2009,벅 등, 2007). 이들의 지속성은 방사선 및 다양한 형태의 화학 요법을 통해 인간 고형 종양의 용적을 성공적으로 줄인 후 거의 불가피하게 질병이 재발하는 것을 설명하는 데 도움이 될 수 있습니다. 실제로, CSC는 특정 형태의 종양 휴면기의 기초가 되는 것으로 입증될 수 있으며, 이에 따라 잠복암 세포는 수술적 절제나 방사선/화학요법 후 수년 또는 심지어 수십 년 동안 지속되다가 갑자기 폭발하여 생명을 위협하는 질병을 발생시킵니다. 따라서 CSC는 치료적 살해에 대한 저항력이 더 강하고 동시에 치료가 중단되면 종양을 재생하는 능력을 부여한다는 점에서 이중 위협을 나타낼 수 있습니다.
CSC 상태에 내재된 이러한 표현형 가소성은 또한 다양한 방식으로 전반적인 종양 성장을 지원하는 종양 내에서 기능적으로 구별되는 하위 집단의 형성을 가능하게 할 수 있습니다. 예를 들어, EMT는 상피 암종 세포를 일부 종양에서 암 관련 섬유아세포(CAF)의 역할을 잘 수행할 수 있는 중간엽 섬유아세포 유사 암세포로 전환할 수 있습니다. 놀랍게도, 몇몇 최근 보고서에서는 교모세포종 세포(또는 이와 관련된 CSC 하위 집단)가 종양 관련 신생혈관을 형성할 때 진정한 숙주 유래 내피 세포를 대체할 수 있는 내피 유사 세포로 전환분화할 수 있는 능력이 문서화되어 있습니다. (소다 외, 2011,엘 할라니 외, 2010,리치-비티아니 외, 2010년,왕 외, 2010). 이와 같은 관찰은 특정 종양이 기능을 제공하기 위해 모집된 숙주 세포에 의존하기보다는 자신의 암세포 중 일부가 다양한 유형의 변태를 거쳐 간질 세포 유형을 생성하도록 유도함으로써 간질 지원을 얻을 수 있음을 나타냅니다.
종양에서 CSC와 생물학적 가소성의 발견은 종양 내 유전적으로 균질한 단일 세포 집단이 그럼에도 불구하고 별개의 분화 상태에 있는 세포의 존재로 인해 표현형적으로 이질적일 수 있음을 나타냅니다. 그러나 표현형 다양성의 똑같이 중요한 원인은 암 진행이 진행됨에 따라 축적되는 종양 내의 유전적 이질성에서 파생될 수 있습니다. 따라서 종양 진행의 후기 단계에서 작용하는 유전적 불안정성의 증가는 다윈의 선택 과정을 능가하는 만연한 유전적 다양성을 촉진하여 제거될 수 있는 것보다 훨씬 더 빠르게 유전적으로 구별되는 하위 집단을 생성할 수 있습니다.
이러한 생각은 최근 DNA(및 RNA) 서열 분석 기술의 주요 발전으로 인해 실용적이게 된 종양 세포 게놈의 심층적인 서열 분석에 의해 점점 더 뒷받침되고 있습니다. 따라서 동일한 종양의 서로 다른 부분에서 미세해부된 암세포의 게놈 서열을 분석합니다(야치다 외, 2010)은 눈에 띄는 종양 내 유전적 이질성을 밝혀냈습니다. 이러한 유전적 다양성 중 일부는 개별 인간 종양 내에서 오랫동안 인식되어 온 조직학적 이질성에 반영될 수 있습니다. 대안적으로, 이러한 유전적 다양화는 기능적 전문화를 가능하게 하여 뚜렷하고 보완적인 능력에 기여하는 암세포의 하위 집단을 생성할 수 있으며, 이는 위에서 설명한 대로 전체 종양 성장의 공통 이점을 얻게 됩니다.
내피 세포
종양 내의 세포 이질성의 대부분은 간질 구획에서 발견됩니다. 간질 구성 요소 중 눈에 띄는 것은 종양 관련 혈관계를 형성하는 세포입니다. 동맥, 정맥, 모세혈관을 구성하는 내피세포의 발달, 분화, 항상성 메커니즘은 2000년에 이미 잘 알려져 있었습니다. 새로운 혈관을 생성할 수 있게 해주는 세포 생물학적 프로그램입니다(위 참조). 지난 10년 동안 내피 세포(예: Notch, Neuropilin, Robo 및 Eph-A/B)에 의해 표시되는 신호 전달 수용체의 리간드와 관련된 상호 연결된 신호 전달 경로 네트워크가 이미 유명한 VEGF인 안지오포이에틴, 그리고 FGF 신호. (파스콸레, 2010,아흐메드와 빅넬, 2009,데자나 외, 2009,카르멜리에와 제인, 2000).
다른 연구 방법으로는 종양 관련 내피 세포의 독특한 유전자 발현 프로파일을 밝히고 정상 세포와 종양 내피 세포의 내강 표면에 표시되는 세포 표면 마커를 식별하는 것입니다. (Nagy 외, 2010,루오슬라티 외, 2010,루오슬라티, 2002). 신호 전달, 전사체 프로필 및 혈관 “우편번호”의 차이는 정상 내피 세포가 종양 관련 내피 세포로 전환되는 과정을 이해하는 데 중요한 것으로 입증될 것입니다. 이러한 지식은 종양 관련 내피 세포를 선택적으로 표적으로 삼기 위해 이러한 차이점을 활용하는 새로운 치료법을 개발할 수 있는 기회로 이어질 수 있습니다.
림프관을 형성하는 세포는 일반 순환계의 내피 세포와 밀접하게 관련되어 있습니다. (타멜라와 알리탈로, 2010). 종양 관련 간질에서의 역할, 특히 종양 성장을 지원하는 역할은 잘 알려져 있지 않습니다. 실제로 고형 종양 내의 간질압이 높기 때문에 종양 내 림프관은 일반적으로 붕괴되고 기능하지 않습니다. 그러나 이와 대조적으로 종양 주변과 암세포가 침입하는 인접한 정상 조직에는 기능적이고 활발하게 성장하는(“림프관 형성”) 림프관이 있는 경우가 많습니다. 이러한 관련 림프관은 다양한 암 유형에서 흔히 관찰되는 배수 림프절에 전이가 파종되는 통로 역할을 할 가능성이 높습니다.
혈관주위세포
앞서 언급한 바와 같이, 혈관주위세포는 혈관의 내피관을 감싸는 손가락 모양의 돌기가 있는 특수한 중간엽 세포 유형(평활근 세포와 관련)을 나타냅니다. 정상 조직에서 혈관주위세포는 일반적으로 정지 상태인 내피세포에 측분비 지지 신호를 제공하는 것으로 알려져 있습니다. 예를 들어, 혈관주위세포에서 분비된 Ang-1은 내피 세포 표면에 발현된 Tie2 수용체가 수신하는 항증식 안정화 신호를 전달합니다. 일부 혈관주위세포는 또한 내피 항상성에서 영양 기능을 하는 낮은 수준의 VEGF를 생성합니다. (Gaengel 외., 2009,버거와 송, 2005). 혈관주위세포는 또한 내피세포와 협력하여 혈관주위세포와 내피세포를 고정하고 혈관벽이 혈류의 정수압을 견딜 수 있도록 돕는 혈관 기저막을 합성합니다.
혈관주위세포의 모집과 결합에 대한 유전적, 약리학적 교란은 종양 내피세포를 지지하는 데 있어 이들 세포의 기능적 중요성을 입증했습니다. (피에트라와 오스트만, 2010,Gaengel 외., 2009,버거와 송, 2005). 예를 들어, 종양 혈관주위세포와 골수 유래 혈관주위세포 전구체에 의해 발현되는 PDGF 수용체를 통한 신호전달의 약리학적 억제는 종양 혈관의 혈관주위세포 범위를 감소시키고 결과적으로 혈관 완전성과 기능을 불안정하게 만듭니다. (피에트라와 오스트만, 2010,라자 외, 2010,Gaengel 외., 2009); 흥미롭게도 이와 대조적으로 정상 혈관의 혈관주위세포는 그러한 약리학적 붕괴를 일으키지 않으며 이는 정상 정지 및 종양 혈관계 조절의 차이에 대한 또 다른 예를 제공합니다. 아직 완전히 입증되지 않은 흥미로운 가설은 혈관계의 혈관주위세포 범위가 열악한 종양이 암세포가 순환계로 침입하는 경향이 더 많아 후속 혈행성 파종을 가능하게 한다는 것입니다. (라자 외, 2010,게르하르트와 셈브, 2008).
면역 염증 세포
위에서도 논의한 바와 같이, 면역 체계의 침윤 세포는 점점 더 종양의 일반적인 구성 요소로 받아들여지고 있습니다. 이러한 염증 세포는 서로 상충되는 방식으로 작동합니다. 종양 길항 및 종양 촉진 백혈구 모두가 전부는 아니더라도 대부분의 종양 병변에서 다양한 비율로 발견될 수 있습니다. 종양 길항 CTL과 NK 세포의 존재는 놀라운 일이 아니지만 기능적으로 특징적인 능력을 향상시키는 면역 세포의 보급은 크게 예상하지 못한 것입니다. 1990년대 후반에 면역계 세포에 의한 신생물 조직의 침윤이 아마도 반직관적으로 종양 진행을 촉진한다는 증거가 축적되기 시작했습니다. 이러한 연구는 만성 염증 부위와 종양 형성의 연관성에 대한 개념적 뿌리를 추적했습니다. (셰퍼와 베르너, 2008:드보르작, 1986). 정상적인 상처 치유 및 감염과 싸우는 과정에서 면역 염증 세포는 일시적으로 나타났다가 사라지는 반면, 만성 염증 부위에서는 면역 염증 세포의 존재가 섬유증, 이상 혈관신생 및 신생물을 비롯한 다양한 조직 병리와 관련되어 있습니다. (그리벤니코프 외, 2010,카린 외, 2006).
지난 10년 동안 다양한 면역 세포 유형의 결정 또는 효과기 기능과 관련된 유전자의 조작과 이러한 세포 또는 그 기능의 약리학적 억제제를 통해 이들이 종양 형성을 촉진하는 데 다양하고 중요한 역할을 하는 것으로 나타났습니다. 종양을 촉진하는 염증 세포 목록에는 이제 대식세포 하위 유형, 비만 세포, 호중구뿐만 아니라 T 및 B 림프구도 포함됩니다. (코펠트 외, 2010,DeNardo 외, 2010,Egeblad 등, 2010,요한슨 외, 2008,머독 외, 2008; DePalma 등, 2007). 이러한 연구를 통해 종양 촉진 작용의 효과기 역할을 하는 염증 세포에서 방출되는 신호 분자의 목록이 늘어나고 있습니다. 여기에는 종양 성장 인자 EGF, 혈관 신생 성장 인자 VEGF, 염증 상태를 증폭시키는 FGF2, 케모카인 및 사이토카인과 같은 기타 혈관 신생 인자; 또한, 이들 세포는 MMP-9 및 기타 기질 금속단백분해효소, 시스테인 카텝신 프로테아제 및 헤파라나아제를 포함하여 혈관신생 전 및/또는 침습성 기질 분해 효소를 생산할 수 있습니다. (키안과 폴라드, 2010,머독 외, 2008). 이러한 다양한 효과기의 발현과 일관되게, 종양 침윤 염증 세포는 종양 혈관신생을 유도 및 유지하고, 암세포 증식을 자극하고, 종양 가장자리에서의 존재를 통해 조직 침범을 촉진하고, 종양을 지지하는 것으로 나타났습니다. 암세포의 전이성 전파 및 파종 (코펠트 외, 2010,Egeblad 등, 2010,키안과 폴라드, 2010,만토바니, 2010,조이스와 폴라드, 2009,만토바니 외, 2008,머독 외, 2008; DePalma 등, 2007).
종양 기질에 존재하는 완전히 분화된 면역 세포 외에도 다양한 부분적으로 분화된 골수 전구세포가 종양에서 확인되었습니다. (머독 외, 2008). 이러한 세포는 골수 기원의 순환 세포와 정상 조직과 염증 조직에서 일반적으로 발견되는 분화된 면역 세포 사이의 매개체를 나타냅니다. 중요하게도, 이들 전구세포는 보다 분화된 파생물과 마찬가지로 입증 가능한 종양 촉진 활성을 가지고 있습니다. 특히 흥미로운 것은 종양 침윤성 골수 세포(대식세포 표지자 CD11b와 호중구 표지자 Gr1을 동시에 발현하는 것으로 정의됨)가 CTL 및 NK 세포 활성을 억제하는 것으로 나타났으며 독립적으로 MDSC로 확인되었습니다. (키안과 폴라드, 2010,오스트랜드-로젠버그와 신하, 2009). 이러한 속성은 특정 골수 세포의 모집이 혈관 신생과 종양 진행을 직접적으로 촉진하는 동시에 면역 파괴를 피할 수 있는 수단을 제공함으로써 발생 중인 종양에 두 배로 유익할 수 있는 가능성을 높입니다.
종양 촉진 및 종양 길항 면역 세포의 반직관적 존재는 면역 체계의 다양한 역할을 호출하여 합리화될 수 있습니다. 한편으로, 면역 체계는 적응성 면역 반응을 통해 감염성 물질을 특이적으로 탐지하고 표적으로 삼습니다. 선천성 면역 체계의 세포에 의해. 다른 한편으로, 선천적 면역 체계는 상처 치유와 죽은 세포 및 세포 잔해 제거에 관여합니다. 이러한 전문적인 작업은 염증 세포의 뚜렷한 하위 클래스, 즉 기존의 대식세포 및 호중구(적응 면역 지원에 관여) 클래스와 상처 치유 및 조직에 관여하는 “대체적으로 활성화된” 대식세포, 호중구 및 골수 전구세포의 하위 클래스에 의해 수행됩니다. 집 청소 (Egeblad 등, 2010,만토바니, 2010,키안과 폴라드, 2010,요한슨 외, 2008). 면역 세포의 후자 아형은 상처 치유에 필요한 혈관 신생, 상피 및 간질 성장 인자와 기질 리모델링 효소의 주요 공급원 중 하나이며 종양 진행을 지원하기 위해 모집되고 전복되는 세포입니다. 마찬가지로, B 및 T 림프구의 하위 클래스는 상처 치유 및 종양 촉진 대식세포 및 호중구의 모집, 활성화 및 지속성을 촉진할 수 있습니다.DeNardo 외, 2010,Egeblad 등, 2010,비스와스와 만토바니, 2010년). 물론, B 및 T 림프구의 다른 하위 클래스와 선천성 면역 세포 유형은 입증 가능한 종양 살해 반응을 나타낼 수 있습니다. 종양에서 상충되는 염증 반응 사이의 균형은 예후에 중요한 역할을 할 가능성이 있으며, 아마도 이러한 세포를 종양 파괴 방향으로 방향을 바꾸도록 고안된 치료법에 도움이 될 것입니다.
암 관련 섬유아세포
섬유아세포는 암종의 스펙트럼 전반에 걸쳐 다양한 비율로 발견되며, 많은 경우 종양 간질의 우세한 세포 집단을 구성합니다. “암 관련 섬유아세포”라는 용어는 적어도 두 가지 별개의 세포 유형을 포함합니다: (1) 대부분의 정상 상피 조직을 지지하는 구조적 기초를 생성하는 섬유아세포와 유사한 세포, (2) 근섬유아세포의 생물학적 역할과 특성은 근섬유아세포와 현저하게 다릅니다. 조직 유래 섬유아세포의. 근섬유아세포는 α-평활근 액틴(SMA)의 발현으로 식별할 수 있습니다. 간이나 췌장과 같은 특정 조직에는 상당량의 α-SMA 발현 세포가 포함되어 있지만 대부분의 건강한 상피 조직에서는 드물다. 근섬유모세포는 상처에서 일시적으로 많이 증가하며 만성 염증 부위에서도 발견됩니다.
모집된 근섬유아세포와 정상 조직 유래 섬유아세포의 재프로그래밍된 변종은 종양 표현형, 특히 암세포 증식, 혈관 신생, 침윤 및 전이를 향상시키는 것으로 나타났습니다. 그들의 종양 촉진 활동은 주로 암세포와 혼합된 암 관련 섬유아세포를 쥐에 이식하는 것으로 정의되었으며, 최근에는 종양이 발생하기 쉬운 쥐에서 그 기능의 유전적, 약리학적 교란으로 정의되었습니다. (디라트 외, 2010,피에트라와 오스트만, 2010,라세넨(Räsänen)과 바헤리(Vaheri), 2010년,시모다 외, 2010,칼루리(Kalluri)와 자이스베르그(Zeisberg), 2006년,Bhowmick 외, 2004). 암 관련 섬유아세포는 다양한 세포외 기질 성분을 분비하기 때문에 많은 진행성 암종의 특징인 조직형성 간질의 형성과 관련이 있습니다. 암 관련 섬유아세포의 두 아형이 종양 발병에 기여하는 기능의 전체 스펙트럼은 아직 밝혀지지 않았습니다.
종양 기질의 줄기 및 전구 세포
종양 미세환경을 구성하는 다양한 간질세포 유형은 이러한 세포 유형의 가장 확실한 저장소인 인접한 정상 조직에서 모집될 수 있습니다. 그러나 최근 몇 년 동안 골수는 종양 관련 간질 세포의 주요 공급원으로 점점 더 많이 관여하고 있습니다. (Bergfeld와 DeClerck, 2010,팽과 살벤, 2011,지아시아와 스키파니, 2010,Patenaude 외, 2010,라마냐와 버거, 2006). 중간엽 줄기세포와 전구세포는 골수에서 종양으로 이동하는 것으로 밝혀졌으며, 그곳에서 잘 특성화된 다양한 간질 세포 유형으로 분화될 수 있습니다. 최근에 도착한 이들 중 일부는 미분화되거나 부분적으로 분화된 상태로 남아 더 분화된 자손에게 부족한 기능을 나타낼 수도 있습니다.
간질 세포 유형의 골수 기원은 골수 세포 및 그에 따라 전파된 자손이 녹색 형광 단백질(GFP)과 같은 리포터로 선택적으로 표지된 종양 보유 마우스를 사용하여 입증되었습니다. 면역 염증 세포는 골수에서 유래하는 것으로 오랫동안 알려져 왔지만, 최근에는 골수에서 유래하는 혈관주위세포와 다양한 아형의 암 관련 섬유아세포의 전구세포가 다양한 암 마우스 모델에서 기술되었습니다. (Bergfeld와 DeClerck, 2010,팽과 살벤, 2011,지아시아와 스키파니, 2010,라마냐와 버거, 2006); 종양 혈관신생에 대한 내피 전구 세포의 유병률과 기능적 중요성은 현재 해결되지 않았습니다. (팽과 살벤, 2011,Patenaude 외, 2010). 종합하면, 이러한 다양한 증거는 종양 관련 간질 세포가 기존 간질 세포의 증식, 이웃 정상 조직에서 유래한 국소 줄기/전구 세포의 제자리 분화, 또는 뼈 모집을 통해 성장하는 종양에 공급될 수 있음을 나타냅니다. 골수 유래 줄기/전구 세포.
이형 신호 전달은 종양 미세 환경의 세포를 조율합니다
암세포 생물학을 지배하는 세포내 회로에 대한 묘사(예: 그림 2 )는 종양 내의 종양 세포와 간질 세포 사이의 복잡한 상호 작용과 그들이 집합적으로 세우고 리모델링하는 역동적인 세포외 기질 사이의 복잡한 상호 작용을 도표화한 유사한 다이어그램으로 보완되어야 합니다. (Egeblad 등, 2010,케센브록 외, 2010,피에트라와 오스트만, 2010,폴리악 외, 2009). 대부분의 신호 분자와 경로가 아직 밝혀지지 않았기 때문에 미세 환경 신호 상호 작용 네트워크를 합리적으로 완전하고 그래픽으로 묘사하는 것은 아직 우리의 손이 닿지 않는 곳에 있습니다. 대신 그림 5 (상단) 에서는 그러한 상호 작용에 대한 힌트를 제공합니다. 이러한 잘 확립된 몇 가지 예는 종양 발병에 매우 중요한 매우 복잡한 신호 전달 네트워크를 예시하기 위한 것입니다.
그림 5 악성 진행 중 종양 미세환경의 신호 상호작용
복잡성의 또 다른 차원은 이 단순한 도식에서 표현되지 않습니다. 정상 조직이 고급 악성종양으로 변하는 다단계 과정에서 종양 세포와 그 주위의 간질 세포가 점진적으로 변합니다. 이러한 조직병리학적 진행은 종양 실질과 간질 사이의 이형 신호 전달의 근본적인 변화를 반영해야 합니다.
이러한 단계적 진행은 도 5 에 도시된 바와 같이 신생물 세포와 지지 간질 세포 사이의 앞뒤 상호 작용에 의존할 가능성이 높습니다. 따라서 초기 신생물은 초기 종양전 간질로 조립되는 간질 세포 유형을 모집하고 활성화함으로써 상호작용을 시작하며, 이는 결국 인근 암세포의 신생물 표현형을 강화함으로써 상호 반응합니다. 유전적으로 더 진화할 수 있는 암세포는 다시 간질에 신호를 다시 공급하여 정상 간질 세포를 재프로그래밍하여 신생 종양에 봉사합니다. 궁극적으로 종양 간질에서 발생하는 신호는 암세포가 정상적인 인접 조직을 침범하여 확산되도록 합니다.
이 상호 이형 신호 전달 모델은 다단계 종양 진행의 최종 단계인 전이를 포괄하도록 확장되어야 합니다( 그림 5)., 오른쪽 아래). 원발성 종양에서 방출된 순환 암세포는 그러한 종양의 지지 간질에 의해 생성된 미세환경을 남깁니다. 그러나 먼 기관에 착륙하면 이러한 암세포는 순진하고 완전히 정상적인 조직 미세환경에 직면하게 됩니다. 결과적으로, 원발성 종양 내에 존재하는 동안 표현형을 형성한 많은 이형 신호가 파종 부위에 없을 수 있으며, 이는 파종된 암세포의 성장에 대한 장벽을 구성합니다. 따라서, 원발성 종양에서 다단계 진행을 정의하는 간질세포 상호작용에 대한 상호 암세포의 계승은 이제 파종된 암세포가 새로 발견된 기관 부위에 정착하기 시작함에 따라 먼 조직에서 새로 반복되어야 합니다.
이 논리는 일부 전이 사례에 적용되지만 앞서 언급한 것처럼 특정 조직 미세 환경은 다양한 이유로 이미 새로 파종된 암세포를 지지할 수 있습니다. 이러한 허용 사이트를 “전이성 틈새”라고 합니다(페이나도 외, 2011,코글린과 머레이, 2010). 이 용어에 함축된 개념은 그러한 부위에 파종된 암세포가 이미 부분적으로 이미 존재하기 때문에 지지 기질을 유도하여 시작할 필요가 없다는 개념입니다. 이러한 허용성은 조직 부위에 고유할 수 있습니다(탈매지와 피들러, 2010) 또는 원발 종양에서 방출된 순환 인자에 의해 사전 유도된 경우(페이나도 외, 2011). 유도된 전이성 틈새의 가장 잘 문서화된 구성 요소는 종양 촉진 염증 세포이지만, 다른 세포 유형과 ECM은 다른 전이 상황에서 중요한 역할을 하는 것으로 잘 입증될 수 있습니다.
다단계 종양 발달 과정에서 암세포와 이를 지지하는 간질 사이의 신호 상호 작용이 진화할 가능성은 암 병인의 메커니즘을 완전히 밝히려는 목표를 분명히 복잡하게 만듭니다. 예를 들어, 이러한 현실은 악성 진행을 조율하는 것보다 중요한 규제 네트워크를 도표화하려는 시스템 생물학자에게 과제를 제기합니다. 더욱이, 이러한 동적 변화를 이해하는 것이 원발성 종양과 전이성 종양 모두를 성공적으로 표적으로 삼도록 고안된 새로운 치료법의 개발에 결정적인 역할을 할 것으로 보입니다.
치료적 표적화
인간 암을 치료하기 위한 메커니즘 기반 표적 치료법의 도입은 암 발병 메커니즘에 대한 지난 30년간의 놀라운 연구 발전의 결실 중 하나로 예고되었습니다. 우리는 여기서 개발 중이거나 최근에 임상에 도입된 수많은 치료법을 열거하려고 시도하지 않습니다. 대신, 우리는 특징 원칙에 대한 설명이 어떻게 현재 치료 개발에 정보를 제공하기 시작했고 앞으로도 점점 더 그렇게 될 수 있는지 고려합니다.
빠르게 성장하고 있는 표적 치료제의 무기고는 그림 6 에 제시된 예에 설명된 것처럼 하나 이상의 특징적인 기능에 대한 각각의 효과에 따라 분류될 수 있습니다. 실제로, 이들 약물의 관찰된 효능은 각 경우에 특정 능력의 검증을 나타냅니다. 만약 어떤 능력이 종양의 생물학에 정말로 중요하다면, 그 억제는 종양의 성장과 진행을 손상시켜야 합니다.
그림 6 암 특징의 치료 표적화
우리는 현재까지 개발된 대부분의 특징 표적 항암제들이 특정 기능을 활성화하는 데 어떤 방식으로든 관여하는 특정 분자 표적을 향해 의도적으로 지향되었다는 점에 주목합니다. 이러한 작용의 특이성은 원칙적으로 표적 외 효과가 상대적으로 적고 따라서 비특이적 독성이 적으면서 표적에 대한 억제 활성을 나타내기 때문에 미덕으로 간주되어 왔습니다. 실제로 결과적인 임상 반응은 일반적으로 일시적이었고 거의 불가피한 재발이 뒤따랐습니다.
증가하는 실험적 증거에 의해 뒷받침되는 이 역사에 대한 한 가지 해석은 각 핵심 특징 기능이 부분적으로 중복된 신호 경로에 의해 규제된다는 것입니다. 결과적으로, 종양의 주요 경로 중 하나를 억제하는 표적 치료제는 특징적인 능력을 완전히 차단하지 못할 수 있으며, 일부 암세포는 해당 세포 또는 그 자손이 결국 적용되는 치료법에 의해 부과되는 선택 압력에 적응할 때까지 잔여 기능을 유지하면서 생존할 수 있습니다. 돌연변이, 후생적 재프로그래밍 또는 간질 미세환경의 리모델링을 통해 달성될 수 있는 이러한 적응은 기능적 능력을 재설정하여 새로운 종양 성장과 임상적 재발을 허용할 수 있습니다. 특정 특징을 지원하는 병렬 신호 전달 경로의 수가 제한되어야 한다는 점을 고려하면,
치료에 반응하여 암세포는 특정 기능에 대한 의존도를 감소시켜 다른 기능에 더 의존하게 될 수도 있습니다. 이는 획득된 약물 저항의 매우 다른 형태를 나타냅니다. 이 개념은 항혈관신생 치료법에 대한 예상치 못한 반응의 최근 발견에 의해 예시됩니다. 일부 사람들은 혈관신생을 효과적으로 억제하면 종양을 휴면 상태로 만들고 심지어 종양을 용해시킬 수도 있다고 예상했습니다. (포크맨과 칼루리, 2004). 대신, 항혈관신생 치료법에 대한 임상적 반응은 일시적인 것으로 밝혀졌습니다. (아잠 외, 2010,에보스 외, 2009,버거와 하나한, 2008).
강력한 혈관신생 억제제가 이러한 특징적인 능력을 억제하는 데 성공한 특정 전임상 모델에서 종양은 지속적인 혈관신생에 대한 의존성에서 대신 다른 혈관신생, 즉 침습성과 전이의 활성을 높이는 방향으로 적응하고 전환합니다. (아잠 외, 2010:에보스 외, 2009,버거와 하나한, 2008). 근처 조직을 침범함으로써 처음에는 저산소 상태의 암세포가 정상적이고 기존의 조직 혈관계에 접근할 수 있는 것으로 보입니다. 이러한 적응/회피 저항에 대한 초기 임상적 검증은 인간 교모세포종을 항혈관신생 요법으로 치료할 때 나타나는 침습 및 국소 전이의 증가에서 명백합니다. (엘리스와 리어던, 2009,Norden 외, 2009,Verhoeff 등, 2009). 이 교훈을 다른 인간 암에 적용할 수 있는지는 아직 확립되지 않았습니다.
다른 특징 특성에 따른 유사한 적응 변화는 또한 유사한 특징 표적 치료법의 효능을 제한할 수도 있습니다. 예를 들어, 세포사멸을 유도하는 약물을 사용하면 암세포가 유사 분열 신호를 과도하게 활성화하여 그러한 치료로 인해 유발된 초기 감소를 보상할 수 있습니다. 이러한 고려 사항은 약물 개발과 치료 프로토콜 설계가 기능적으로 분리된 특징 능력과 각 기능을 지원하는 데 관련된 다양한 생화학적 경로의 개념을 통합함으로써 이익을 얻을 수 있음을 시사합니다. 따라서 특히 우리는 다중 핵심 및 새로운 특징 기능과 활성화 특성의 선택적 공동 타겟팅을 구상할 수 있습니다( 그림 6) 메커니즘 기반 조합을 통해 인간 암에 대한 보다 효과적이고 내구성 있는 치료법이 탄생할 것입니다.
결론 및 미래 비전
우리는 여기에서 암의 복잡한 생물학을 이해하는 데 유용한 개념적 틀을 제공한 암 특징의 개념을 재검토하고, 개선하고, 확장하려고 노력했습니다. 암의 특징인 6가지 획득 능력은 대부분의 암 형태의 필수 구성 요소로서 시간의 시험을 견뎌왔습니다. 지난 10년간 놀라운 개념적 발전을 이어가면서 이러한 조직 원칙은 가까운 미래에 더욱 정교해질 것입니다.
앞으로 우리는 침입과 전이에 대한 이해에 있어서 향후 10년 동안 상당한 발전을 이룰 것으로 예상합니다. 마찬가지로, 이러한 대사 재프로그래밍이 만성적으로 지속되는 증식의 핵심 특징과 분리될 수 있는 개별 기능인지 여부에 대한 해결을 포함하여 악성 성장에서 호기성 해당작용의 역할이 밝혀질 것입니다. 우리는 면역 감시가 사실상 모든 종양이 우회해야 하는 장벽인지, 아니면 특히 면역원성인 일부 종양의 특이성인지에 대해 여전히 혼란스러워하고 있습니다. 이 문제 역시 어떤 식으로든 해결될 것입니다.
그러나 다른 영역은 현재 빠르게 변화하고 있습니다. 최근 몇 년 동안 염색질 변형을 통해 전사를 제어하는 정교한 분자 메커니즘이 밝혀졌으며 특정 특징 능력을 획득하는 동안 염색질 구성의 특정 변화가 발생한다는 단서가 있습니다. (베르다스코와 에스텔러, 2010). 기능적으로 중요한 후생유전학적 변화는 암세포뿐만 아니라 종양 관련 간질의 변형된 세포에서도 요인이 될 가능성이 높습니다. 이러한 후생적 메커니즘의 해명이 특징적인 능력을 획득하는 수단에 대한 우리의 전반적인 이해를 실질적으로 변화시킬 것인지, 아니면 단순히 이를 지배하는 것으로 이미 알려진 규제 회로에 추가적인 세부 사항을 추가할 것인지는 현재로서는 불분명합니다.
마찬가지로, 수백 가지의 서로 다른 조절 마이크로RNA의 발견은 이미 건강과 질병에서 작동하는 유전자 제어 메커니즘에 대한 우리의 이해에 중대한 변화를 가져왔습니다. 지금까지 수십 개의 마이크로RNA가 다양한 종양 표현형과 관련되어 있습니다.Garzon 외, 2010), 그러나 이는 우리 세포에 존재하고 다양한 형태의 암에서 발현이 변경된 것으로 알려진 수백 개의 마이크로RNA의 기능이 완전한 미스터리로 남아 있기 때문에 실제 복잡성의 표면을 긁을 뿐입니다. 여기서도 우리는 미래의 진보가 암의 병인 메커니즘에 대한 우리의 이해에 근본적인 변화를 가져올 것인지, 아니면 이미 계획된 정교한 규제 회로에 세부 사항을 추가할 것인지에 대해 불분명합니다.
마지막으로, 서로 다른 형태와 점진적으로 악성화되는 암 단계를 생성하기 위해 조립되고 협력하는 여러 별개의 세포 유형 간의 이형 상호작용에 대한 회로도는 현재 초보적입니다. 앞으로 10년 안에 우리는 종양 내의 다양한 세포 사이의 상호소통을 설명하는 신호 회로가 훨씬 더 자세하고 명확하게 차트로 작성되어 현재 지식을 능가할 것으로 예상합니다. 그리고 이전과 마찬가지로 (하나한과 와인버그, 2000), 우리는 암 연구를 점점 더 논리적인 과학으로 예측하고 있으며, 여기서 무수한 표현형 복잡성은 기본 구성 원리의 작은 집합을 나타냅니다.
번역 후 변형(PTM)은 정상 세포와 암세포 모두에서 세포 신호 전달 및 생리학을 조절하는 데 중요한 역할을 합니다. 질량 분석법의 발전으로 높은 처리량, 정확하고 민감한 PTM 수준 측정이 가능해 PTM 수준의 역할, 보급 및 누화를 더 잘 이해할 수 있습니다. 여기에서 우리는 11가지 암 유형에 걸쳐 PTM 프로파일을 가진 1,110명의 환자로부터 가장 큰 단백질 유전학 데이터 컬렉션을 분석합니다(국립 암 연구소의 임상 단백질 종양 분석 컨소시엄(CPTAC)의 10명). 우리의 연구는 특징적인 암 과정과 관련된 단백질 아세틸화 및 인산화 변화의 범암 패턴을 보여줍니다. 이러한 패턴은 인산화에 의해 DNA 복구가 조절되지 않는 암을 포함하여 다양한 암 유형의 종양 하위 집합을 나타냅니다. 아세틸화에 의한 면역 반응과 관련된 대사 조절의 변화, 아세틸화와 인산화 사이의 누화에 의한 키나제 특이성 영향, 히스톤 조절의 변형. 전반적으로 이 리소스는 PTM이 적용되는 풍부한 생물학을 강조하고 잠재적인 새로운 치료 방법을 제시합니다.
소개
종양에 대한 체계적인 유전체학 기반 연구는 종양 생물학에 대한 이해에 혁명을 일으켰습니다. 환자 치료에 큰 영향을 미쳤습니다. 그러나 많은 암은 여전히 효과적인 치료법이 부족하거나 특성이 제대로 규명되지 않아 복잡한 생물학과 분자 및 표현형 이질성이 강조됩니다. 시료 처리 및 액체 크로마토그래피-탠덤 질량 분석법(LC-MS/MS)의 최근 발전으로 단백질 수준과 번역 후 변형(PTM)을 대규모로 정량화할 수 있게 되었습니다.
CPTAC(Clinical Proteomic Tumor Analysis Consortium)의 공동 노력으로 개별 암 유형에 대한 대규모 단백질유전체 데이터 세트가 생성되었습니다. 이 연구에는 모두 PTM이 포함되었으며 분자적 특징과 표현형 결과 사이의 격차를 해소하여 잠재적인 치료 취약성을 지닌 새로운 암 하위 유형을 식별하기 시작했습니다. 이러한 발전과 PTM이 세포 신호 전달을 조절하고 미세 조정하는 데 중요한 역할을 하고 있음에도 불구하고, 공유 패턴, PTM 간의 누화(예: 인산화, 아세틸화 등), 여러 PTM이 규제 네트워크를 형성하는 방식, 특히 암 유형 전반에 걸쳐 잘 이해되지 않은 상태입니다.
이전의 범암 게놈 연구에서는 다양한 암 유형에 걸쳐 재발성 유전자 및 경로 변경을 조사하면 암을 유발하는 근본적인 분자 현상에 대한 이해를 높일 수 있음이 입증되었습니다. 여기에서 우리는 게놈 연구를 확장하고 보완하기 위해 여러 암에서 변경되는 일반적인 번역 후 조절 메커니즘을 조사하기 위해 암 유형 전반에 걸쳐 공유되고 다양한 PTM 패턴을 식별하기 시작했습니다. 이를 위해 우리는 완전한 게놈, 전사체, 단백질체 및 PTM(인산화 및 아세틸화) 데이터를 갖춘 1,110명의 치료 경험이 없는 환자의 샘플을 포함하는 11개 연구의 데이터를 사용하여 조화된 범암 코호트를 생성했습니다(그림 1A ). 이를 통해 개별 연구의 제한된 표본 크기(환자 39~140명)로 인해 단일 코호트에서 식별할 수 없는 패턴을 검색할 수 있었습니다. 암 전체에 걸쳐 공유되고 조직 독립적인 패턴에 초점을 맞추기 위해 우리는 조화 과정의 일부로 각 데이터 유형의 조직별 효과를 회귀 분석했습니다.
우리는 암에서 조절 장애가 있는 것으로 알려진 (1) 특징적인 경로에 대한 분석을 집중했습니다. PTM에 의해 엄격하게 통제되며, DNA 손상 및 복구 경로, 세포 면역 대사 및 유전자 발현의 히스톤 수준 조절을 포함합니다. (2) 다양한 유형의 PTM 간의 잠재적 누화. PTM은 빠른 것부터 지속적인 것, 장기적인 것까지 다양한 잠재적 규제 효과를 가지고 있습니다. 면역 및 대사 반응에서 PTM의 일시적이고 가역적인 특성은 미세 환경의 변화에 적응하는 데 필요한 빠른 반응을 가능하게 합니다. 반면, 히스톤 변형에 대한 PTM 효과는 세포 프로그램의 장기간 지속되는 조절에 영향을 미칠 수 있습니다. 실제로 암에서 비정상적인 히스톤 아세틸화는 종양 억제 인자를 비활성화하거나 종양 유전자를 활성화할 수 있습니다. DNA 복구 과정에서 인산화는 DNA 복구 단백질의 활성을 조절하는 데 중요한 역할을 합니다. PTM에 초점을 맞춘 분석은 특히 DNA 복구가 부족한 암에서 DNA 복구 환경을 더 잘 특성화할 수 있습니다. 마지막으로, 세린/트레오닌 인산화와 라이신 아세틸화는 진핵생물에서 가장 널리 퍼져 있고 보존된 PTM 중 하나입니다. 현재까지 대부분의 연구는 단일 PTM 유형이 세포 과정을 어떻게 조절할 수 있는지에 초점을 맞추었지만, 단백질이 여러 PTM 유형을 가지고 있다는 인식은 이들이 함께 작용하여 복잡한 규제 효과를 공동으로 나타낼 수 있음을 시사하며 그 중 대부분은 아직 탐구되지 않은 상태입니다.
전반적으로, 이것은 아세틸화와 인산화의 광범위한 조절과 암 유형 전반에 걸친 공유 패턴을 자세히 설명하는 최초의 범암 연구입니다. 함께, 우리의 결과는 추가 실험 검증 후 새로운 약물 표적을 식별하거나 암 생물학에 영향을 미치는 새로운 방법을 제안할 수 있는 암의 PTM 관리 프로세스에 관한 가설을 탐색하고 생성하는 풍부한 리소스로 구성됩니다.
결과(원문에서 확인 가능함)
Pan-cancer 데이터 세트 개요
범암 PTM 환경
DNA 복구 결핍 종양에서 PTM 조절 장애의 메커니즘
대사 경로의 PTM 조절은 종양 관련 면역 반응에 영향을 미칩니다
암 관련 유전자에서 PTM에 의한 히스톤 조절의 변화
암에서 단백질 인산화와 아세틸화 사이의 누화
논의
PTM은 신호 전달의 핵심 조절자이며 다른 많은 필수 기능 중에서 단백질 간 상호 작용, 단백질 안전성 및 위치 파악에 중요한 역할을 합니다. 이 연구에서 우리는 11가지 암 유형에 걸쳐 PTM을 종합적으로 조사하고 알려진 암 특징 프로세스(1) DNA 복구, (2) 면역 반응, (3) 대사, (4) 히스톤 조절 및 (5)에 대한 PTM의 기여를 강조했습니다. 우리는 암 유형 전반에 걸쳐 이러한 과정과 PTM 패턴의 공통점과 중요한 차이점을 지적했습니다. 이 풍부한 리소스를 통해 여기서 설명하는 PTM 작업을 넘어 암 유형 전반에 걸쳐 PTM에 대한 추가 조사가 가능해집니다.
HRD 및 MMRD와 같은 DNA 복구 결함은 종양 발달 전반에 걸쳐 체세포 돌연변이 패턴을 생성하여 주어진 복구 결함의 증거를 제공합니다. 중요한 것은 이러한 돌연변이 시그니처가 복구 경로의 현재 활동을 반드시 반영하는 것은 아니며, 이는 DNA 복구 유전자를 표적으로 하는 치료법(예: PARP 및 POLQ 억제제 등)에 대한 반응의 변화를 이해하는 데 적합할 수 있다는 것입니다. DNA 복구가 부족한 암에 대한 심층 분석은 인산화 중심 분석이 게놈 및 전사체 수준에서 감지할 수 없는 정보 패턴을 밝히고 특성화하는 능력을 강조했습니다. HRD 클러스터 분석을 통해 우리는 DNA 복구 단백질의 인산화의 중요한 차이가 저산소증의 심각도와 밀접한 관련이 있음을 발견했습니다. 우리는 만성 저산소증이 있는 HRD 종양에서 PARP1을 포함한 여러 DNA 복구 단백질의 활성이 감소하여 잠재적으로 PARP 억제제에 대한 반응에 영향을 미치는 것을 발견했습니다. MMRD 종양에 대한 우리의 단백질유전체학적 분석은 재발성 RAD50 미세부수체 삽입결실과 DSB 감지 및 신호 전달에 중요한 MRN 복합체의 세 가지 단백질 풍부도가 심하게 감소한 것과 연결되었습니다. 부위별 인산화 분석을 통해 우리는 DSB 복구 기능 장애에 대한 추가 증거를 확인했으며 이는 MMRD(즉, MSI) 암 치료법 개발을 위한 추가 방법을 제공할 수 있습니다.
일반적으로 면역 반응은 숙주가 직면한 특정 위협에 대한 각 반응을 맞춤화하기 위해 엄격하게 규제되므로 PTM이 달성할 수 있는 신속한 규제 변화가 필요합니다. 마찬가지로 세포 대사에도 같은 유연성이 필요하므로 PTM은 면역 반응과 대사 반응을 모두 조절하는 데 필수적인 역할을 합니다. 예를 들어, 이 두 과정은 FA 효소에 대한 PTM에 의한 지질 대사의 암세포 조절이 면역 반응에 영향을 미칠 수 있다는 증가하는 증거와 연결될 수 있습니다. 이 연구에서 우리는 아세틸화에 의해 유도되는 다양한 대사 표현형을 갖는 4개의 발현 기반 면역 클러스터를 확인했습니다. CLUMPS-PTM은 해당과정 관련 단백질인 ALDOA를 강조했습니다. 이 단백질은 ALDOA 활성 증가와 관련된 면역 핫 하위 유형의 변경된 인산화 및 아세틸화 부위의 중요한 클러스터를 모두 가지고 있습니다. 생쥐에서 ALDOA를 억제하면 폐 전이가 감소하고 생존 기간이 연장됩니다. Immuno-cool 하위 유형은 IFNγ 발현 감소와 밀접한 상관관계가 있는 FA 대사 활동의 증가를 보여주었는데, 이는 면역 억제에서 FA의 중요한 역할을 암시합니다. 최근 연구에 따르면 급성 골수성 백혈병(AML)에서 FA 산화를 억제하면 내성이 생긴 세포에서 베네토클락스 및 아자시티딘에 대한 민감성을 회복할 수 있는 것으로 나타났습니다. 이러한 결과는 암에서 지질 대사를 표적으로 삼으면 종양 세포가 더 높은 수준의 에너지를 생산하는 능력을 감소시킬 뿐만 아니라 면역 세포 침투 및 활성화에 더 도움이 되는 종양 미세 환경을 촉진할 수 있음을 강조합니다. 또한, 우리는 잠재적으로 아세틸화와 에너지 생성을 위해 세포에서 사용되는 아세틸 CoA의 가용성 감소로 인해 전사 활성 대사 경로와 히스톤 아세틸화 감소 사이의 연관성을 확인했습니다. 이 규정을 더 자세히 조사하려면 추가 연구가 필요할 것입니다.
마지막으로, 우리는 The Kinase Library를 사용하여 아세틸화와 인산화 사이의 누화에 대한 포괄적인 분석을 수행했습니다. 이 분석은 대부분의 세린/트레오닌 키나제가 인산화 부위에 근접한 아세틸화 리신을 선호하지 않음을 보여 주었으며(이웃 아세틸-/인산 부위의 유의하게 음의 상관관계가 있는 쌍으로 표시됨) 누화에 대한 책임이 있는 키나제를 예측할 수 있게 해줍니다.
요약하면, PTM은 종양 세포의 적응과 세포 내 및 환경 변화에 대한 반응의 필수적인 부분입니다. 암의 시작과 진행으로 이어지는 PTM 관리 프로세스에 대한 더 깊은 이해는 새로운 치료 목표를 밝히고, 기존 치료법에 대한 반응의 바이오마커를 식별하고, 암 생물학에 대한 지식을 확장할 수 있는 잠재력을 가지고 있습니다.
연구의 한계
게놈 기반 범암 연구는 새로운 암 유발 유전자와 공유된 조절 장애 경로를 발견하고 실행 가능한 치료 목표를 식별하는 데 매우 귀중한 자원임이 입증되었습니다. 우리의 단백질유전체학 범암 연구는 11개 종양 유형에 걸쳐 1,110개의 샘플로 제한되었으며, 더 많은 사례와 더 많은 암 유형을 포함하는 대규모 연구가 암의 근본적인 단백질유전체학 메커니즘을 식별하는 힘을 높일 것으로 기대합니다. 이 연구에 설명된 분석은 모두 대량 종양 물질을 기반으로 합니다. 단일 세포 및 공간 전사체 분석과 유사하게, 단일 세포 단백질체학, 공간 단백질체학, 레이저 포착 현미해부를 포함한 기술 종양의 이질성과 특정 세포 유형이 암에 미치는 영향에 대한 더욱 귀중한 통찰력을 제공할 가능성이 큽니다. PTM 누화에 대한 포괄적인 PTM 중심 연구 및 분석이 새로운 분야이지만 몇 가지 단점은 주목할 가치가 있습니다. (1) 현재 질량 분석법 분석은 PTM 간의 누화 분석을 수행하는 능력을 제한하는 상대적으로 높은 위음성 비율을 가지고 있습니다. (2) PTM 간의 누화 관계를 완전히 설정하려면 2개 이상의 PTM을 동시에 감지하기 위한 이중 MS 검색이 필요합니다. (3) 인산화 데이터베이스 및 키나제 예측 도구가 점점 늘어나고 있지만, 병렬적인 포괄적인 아세틸화 데이터베이스 및 도구는 현재 부족하며 본 연구에서 보고된 많은 아세틸화 부위의 기능적 효과는 아직 탐구되지 않고 있습니다.